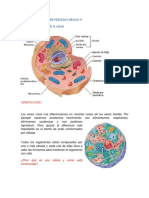
También podría gustarte
- Teórica II BiocelDocumento10 páginasTeórica II BiocelcatalinaAún no hay calificaciones
- BiocelDocumento14 páginasBiocelprofesor manuelAún no hay calificaciones
- Capitulo X. CitoesqueletoDocumento8 páginasCapitulo X. CitoesqueletoCamila Paulina Aravena SotoAún no hay calificaciones
- 16 - Ayudas - CitoesqueletoDocumento9 páginas16 - Ayudas - CitoesqueletoMario AlzamoraAún no hay calificaciones
- Desgrabado de Video - Citoesqueleto IIDocumento6 páginasDesgrabado de Video - Citoesqueleto IIAgustina FernandezAún no hay calificaciones
- CitoesqueletoDocumento15 páginasCitoesqueletoAlba Macipe FeitoAún no hay calificaciones
- BC 8 NecroticaenfmedDocumento6 páginasBC 8 NecroticaenfmedCamilaAún no hay calificaciones
- Citoesqueleto y CitosolDocumento17 páginasCitoesqueleto y CitosolRosaAún no hay calificaciones
- CitoesqueletoDocumento45 páginasCitoesqueletovalentinAún no hay calificaciones
- Citoesqueleto Clase 2023Documento57 páginasCitoesqueleto Clase 2023Nabila InsfranAún no hay calificaciones
- 9 - Citoesqueleto - Medicina 2021Documento14 páginas9 - Citoesqueleto - Medicina 2021Lomito CompletoAún no hay calificaciones
- CITOESQUELETODocumento29 páginasCITOESQUELETOElisandra Rodriguez GonzalesAún no hay calificaciones
- Biologia Celular y Molecular - HistologiaDocumento10 páginasBiologia Celular y Molecular - Histologiapimenteltamara2Aún no hay calificaciones
- CITOESQUELETODocumento12 páginasCITOESQUELETOpimenteltamara2Aún no hay calificaciones
- CitoesqueletoDocumento5 páginasCitoesqueletocamilaonorioAún no hay calificaciones
- Microfilamentos ExpoDocumento18 páginasMicrofilamentos ExpoKelly Lorena Bustamante RegaladoAún no hay calificaciones
- Tema 5 AcabadoDocumento10 páginasTema 5 AcabadoMagdalena Oñate SerranoAún no hay calificaciones
- F ActinaDocumento10 páginasF ActinaAgustina UriarteAún no hay calificaciones
- Cito EsqueletoDocumento8 páginasCito EsqueletoMarilyn MontecinosAún no hay calificaciones
- Citoesqueleto, ResumenDocumento13 páginasCitoesqueleto, ResumenMKsander GarcíaAún no hay calificaciones
- Cito EsqueletoDocumento7 páginasCito Esqueletonaini honoratoAún no hay calificaciones
- CitoesqueletoDocumento6 páginasCitoesqueletoLoreto Santana SchwarzeneggerAún no hay calificaciones
- Citoesqueleto 2011Documento33 páginasCitoesqueleto 2011Jaqueline Romina GonzálezAún no hay calificaciones
- CITOESQUELETODocumento7 páginasCITOESQUELETOLuana HuertasAún no hay calificaciones
- Taller Resuelto 2do ParcialDocumento16 páginasTaller Resuelto 2do ParcialJERONIMO GOMEZ GIRALDOAún no hay calificaciones
- BIO Tema 7. Citoesqueleto (Parte I)Documento11 páginasBIO Tema 7. Citoesqueleto (Parte I)Carmen LostalAún no hay calificaciones
- Tema 15 - Citoesqueleto IIDocumento6 páginasTema 15 - Citoesqueleto IIPau Ruiz MolinaAún no hay calificaciones
- CitoesqueletoDocumento8 páginasCitoesqueletoMili SassoneAún no hay calificaciones
- S6 - CitoesqueletoDocumento8 páginasS6 - CitoesqueletoJhony Farith Farfán ReáteguiAún no hay calificaciones
- Biologia 7Documento15 páginasBiologia 7Lou-LouAún no hay calificaciones
- BiologíaDocumento3 páginasBiologíaNoe RomoAún no hay calificaciones
- Citoesqueleto Seminario 2Documento18 páginasCitoesqueleto Seminario 2Luu GonzalezAún no hay calificaciones
- CitoesqueletoDocumento5 páginasCitoesqueletoSofia Isabella GuevaraAún no hay calificaciones
- CITOESQUELETODocumento95 páginasCITOESQUELETORafael Angel Mejias IzquierdoAún no hay calificaciones
- CITOESQUELETO Especializaciones RevisiónDocumento75 páginasCITOESQUELETO Especializaciones Revisiónalejandra battiAún no hay calificaciones
- CitoesqueletoDocumento22 páginasCitoesqueletomaria aline rezende de souzaAún no hay calificaciones
- Citoesquelto Resumen Cap12Documento9 páginasCitoesquelto Resumen Cap12claudiaAún no hay calificaciones
- UNIDAD #06-Citosol o Matriz CitoplasmáticaDocumento18 páginasUNIDAD #06-Citosol o Matriz CitoplasmáticaJaqueline Alexias LozanoAún no hay calificaciones
- Biologia - CitoesqueletoDocumento26 páginasBiologia - CitoesqueletoJuanita AcostaAún no hay calificaciones
- UNIDAD 5 CitoesqueletoDocumento58 páginasUNIDAD 5 CitoesqueletoCarolina Bravo BastidasAún no hay calificaciones
- Resumen CitoesqueletoDocumento14 páginasResumen CitoesqueletoAmaury de la CruzAún no hay calificaciones
- Wuolah Free App 1707050491591Documento8 páginasWuolah Free App 1707050491591marinagarciapalomaresAún no hay calificaciones
- Los Filamentos de Actina en La Zona Apical Del Intestino DelgadoDocumento5 páginasLos Filamentos de Actina en La Zona Apical Del Intestino DelgadoYisenia VillegasAún no hay calificaciones
- TEMA 6 BiocelDocumento7 páginasTEMA 6 BiocelevamatiasalfonsoAún no hay calificaciones
- Cito EsqueletoDocumento68 páginasCito EsqueletoDiego PantojaAún no hay calificaciones
- CitoesqueletoDocumento66 páginasCitoesqueletoBetty PostigoAún no hay calificaciones
- Unidad 2 - CitoesqueletoDocumento41 páginasUnidad 2 - CitoesqueletoMajo CalderonAún no hay calificaciones
- Unidad 9 CitoesqueletoDocumento43 páginasUnidad 9 CitoesqueletoAlba Fernández RevueltaAún no hay calificaciones
- 11 Citoesqueleto PDFDocumento48 páginas11 Citoesqueleto PDFIrene CarballoAún no hay calificaciones
- CitoesqueletoDocumento102 páginasCitoesqueletoMarco David Meza SerpaAún no hay calificaciones
- MICROFILAMENTOSDocumento7 páginasMICROFILAMENTOSalmanza_1973Aún no hay calificaciones
- Citoesqueleto y Enfermedades DegenerativasDocumento25 páginasCitoesqueleto y Enfermedades DegenerativasJuan Salas FigueroaAún no hay calificaciones
- Taller Escrito CitoesqueletoDocumento5 páginasTaller Escrito Citoesqueletomanda.gnzlzAún no hay calificaciones
- Resumen Capítulo 17 CitoesqueletoDocumento7 páginasResumen Capítulo 17 CitoesqueletoNorma FrancoAún no hay calificaciones
- Tema 6 BiocelDocumento8 páginasTema 6 Biocelrodriguezmartinezm022Aún no hay calificaciones
- 3.1 Matriz Citoplasma - Tica y CitoesqueletoDocumento33 páginas3.1 Matriz Citoplasma - Tica y CitoesqueletoElvirita Vanessa PereiraAún no hay calificaciones
- Cuestionario Actina y MiosinaDocumento6 páginasCuestionario Actina y MiosinaGenek DiazAún no hay calificaciones
- 11.-Expliquen La Motilidad Extramuscular Y Ocúpese Sobre Las Proteínas de Unión Con La Actina Motilidad ExtramuscularDocumento4 páginas11.-Expliquen La Motilidad Extramuscular Y Ocúpese Sobre Las Proteínas de Unión Con La Actina Motilidad ExtramuscularORIALIZ IRAIDA BOMBILLA LEON0% (1)
- CentríoloDocumento65 páginasCentríoloAlfonso Martin SanchezAún no hay calificaciones
- Inducción Al Estudio de Las VariablesDocumento40 páginasInducción Al Estudio de Las VariablesEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Naturaleza Juridica Derecho RegistralDocumento2 páginasNaturaleza Juridica Derecho RegistralEMILY MARIA GALINDO HERNANDEZ100% (1)
- PRESCRIPCIONDocumento8 páginasPRESCRIPCIONEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Antecedentes Históricos Derecho RegistralDocumento46 páginasAntecedentes Históricos Derecho RegistralEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Modelo Certificacion de Nacimiento GuatemalaDocumento1 páginaModelo Certificacion de Nacimiento GuatemalaEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Resolucion de Tramite PruebaDocumento1 páginaResolucion de Tramite PruebaEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Actuacion de Las Partes 18-8-2020Documento15 páginasActuacion de Las Partes 18-8-2020EMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Hojas de Trabajo Física PDFDocumento60 páginasHojas de Trabajo Física PDFEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Tercer Parcial /2014 Ud Quimica Instrucciones Generales Del ExamenDocumento6 páginasTercer Parcial /2014 Ud Quimica Instrucciones Generales Del ExamenEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Pacto de SumisionDocumento1 páginaPacto de SumisionEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Pesentacion Marco MetodologicoDocumento10 páginasPesentacion Marco MetodologicoEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Ji Cuadrada, 2014 PDFDocumento7 páginasJi Cuadrada, 2014 PDFEMILY MARIA GALINDO HERNANDEZAún no hay calificaciones
- Inserto para Determinacion de Ac. Anti HivDocumento3 páginasInserto para Determinacion de Ac. Anti HivGENDRITZAún no hay calificaciones
- Bio. (02) Ciclo CelularDocumento8 páginasBio. (02) Ciclo CelularRuth PumarrumiAún no hay calificaciones
- Taller CITOESQUELETODocumento6 páginasTaller CITOESQUELETOYuyitoS2714Aún no hay calificaciones
- Contenidos Tercer Periodo Grado 4 LA CELULADocumento13 páginasContenidos Tercer Periodo Grado 4 LA CELULALuz Arifa EcheverryAún no hay calificaciones
- Guia de Laboratorio VirtualDocumento3 páginasGuia de Laboratorio VirtualRonal AnayaAún no hay calificaciones
- Tinción de GramDocumento8 páginasTinción de GramStebanContrerasAún no hay calificaciones
- S8 MiologíaDocumento50 páginasS8 MiologíaJORGE LUIS HUAMANI CALIXTOAún no hay calificaciones
- Citología IDocumento52 páginasCitología IMarco PaucarAún no hay calificaciones
- La CelulaDocumento70 páginasLa CelulaRicardo ClementeAún no hay calificaciones
- La CélulaDocumento17 páginasLa Célulafatycas0% (1)
- Tejido Cartilaginoso y ÓseoDocumento19 páginasTejido Cartilaginoso y ÓseoJunior Mejía PonceAún no hay calificaciones
- Tejido MuscularDocumento14 páginasTejido MuscularAndrea Chacón R / ColombiaAún no hay calificaciones
- La CelulaDocumento6 páginasLa CelulaMelissa Andrea Figueredo MontesAún no hay calificaciones
- Clases de CélulasDocumento2 páginasClases de CélulasOswa QuejAún no hay calificaciones
- 11ava PRACTICA PATO GEN - PIEL-Endocri San BorjaDocumento37 páginas11ava PRACTICA PATO GEN - PIEL-Endocri San Borjamax CamacAún no hay calificaciones
- Hueso y CementoDocumento4 páginasHueso y CementoMelanie LozadaAún no hay calificaciones
- Acordeón Histología IDocumento8 páginasAcordeón Histología Ichrised_xAún no hay calificaciones
- Tejido ÓseoDocumento4 páginasTejido ÓseoBarryMedAún no hay calificaciones
- Tarea Domiciliaria CelulaDocumento5 páginasTarea Domiciliaria CelulaEdDy Olivares Gomez0% (1)
- Fisiologia de HuesoDocumento21 páginasFisiologia de HuesoMaritzaAún no hay calificaciones
- Preparacion TejidosDocumento47 páginasPreparacion TejidosFranz mendelevilAún no hay calificaciones
- Hoja de Trabajo Laboratorio 3, Estudiantes 2015Documento6 páginasHoja de Trabajo Laboratorio 3, Estudiantes 2015Alexandra EscobarAún no hay calificaciones
- Tejido ConectivoDocumento1 páginaTejido ConectivoMARICIELOAún no hay calificaciones
- Célula VegetalDocumento2 páginasCélula VegetalEdwin ramirezAún no hay calificaciones
- Bio S4 2020 IiDocumento20 páginasBio S4 2020 IijosssAún no hay calificaciones
- CLAVE Morfo I Temario 1Documento6 páginasCLAVE Morfo I Temario 1Javier Diaz CastroAún no hay calificaciones
- Sopa de LetrasDocumento2 páginasSopa de LetrasMiguel StrikAún no hay calificaciones
- TerminologiaDocumento14 páginasTerminologiamanuelAún no hay calificaciones
- Cuadernillo 2Documento31 páginasCuadernillo 2Fredy Alba100% (1)
- EndosimbiosisDocumento4 páginasEndosimbiosisdiego fernando lopezAún no hay calificaciones