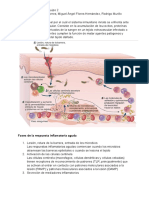
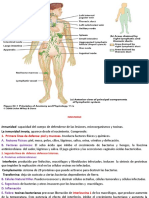
También podría gustarte
- InflamaciónDocumento9 páginasInflamaciónIGNACIO SALVADOR FERNANDEZ GIRONAún no hay calificaciones
- INMUNOLOGDocumento27 páginasINMUNOLOGortiz1081Aún no hay calificaciones
- Inmunidad Innata y AdaptativaDocumento6 páginasInmunidad Innata y AdaptativaBella Jossie100% (1)
- Inmuno AgueDocumento57 páginasInmuno Aguemarinachocolateblanco.negroAún no hay calificaciones
- 2.1 Inflamacion 2023-2Documento113 páginas2.1 Inflamacion 2023-2Maria Fernanda Tordoya ChavezAún no hay calificaciones
- Migracion de Los Leucocitos A Los TejidosDocumento6 páginasMigracion de Los Leucocitos A Los TejidosVíctor ArticaAún no hay calificaciones
- Barrera SecundariaDocumento5 páginasBarrera SecundariaDereck AlayoAún no hay calificaciones
- INFLAMACIONDocumento23 páginasINFLAMACIONSandra chopinAún no hay calificaciones
- Inmuno Temas 1-4Documento34 páginasInmuno Temas 1-4marinachocolateblanco.negroAún no hay calificaciones
- Mecanismos Efectores de La Respuesta Inmune InnataDocumento7 páginasMecanismos Efectores de La Respuesta Inmune InnataEmilis MadridAún no hay calificaciones
- Bases 1Documento9 páginasBases 1Barbara AzevedoAún no hay calificaciones
- Fisio Tema 30Documento8 páginasFisio Tema 30lucisAún no hay calificaciones
- Inflamacion y Reparacion - Patologia - RobbinsDocumento14 páginasInflamacion y Reparacion - Patologia - Robbinslidia chambi zelaAún no hay calificaciones
- Informe de InflamacionDocumento8 páginasInforme de InflamacionflorcitaAún no hay calificaciones
- Respuesta InflamatoriaDocumento7 páginasRespuesta InflamatoriaAngie JiménezAún no hay calificaciones
- Clase 1 - Inflamacion Conceptos GeneralesDocumento8 páginasClase 1 - Inflamacion Conceptos GeneralesDomiJojotAún no hay calificaciones
- Resumen Inflamacion Patologia RobbinsDocumento7 páginasResumen Inflamacion Patologia RobbinsLorena Vera UbillúsAún no hay calificaciones
- Tema 2 - Inflamacion y FiebreDocumento5 páginasTema 2 - Inflamacion y FiebreRodrigo LeesAún no hay calificaciones
- Plan de Clase MFPH I. AO 6Documento11 páginasPlan de Clase MFPH I. AO 6AngelAún no hay calificaciones
- Fagocitosis InflamaciónDocumento10 páginasFagocitosis InflamaciónFelipe Tang CamposAún no hay calificaciones
- InflamaciónDocumento8 páginasInflamaciónJhon HortealesAún no hay calificaciones
- Fisiopatologia de La InflamaciónDocumento27 páginasFisiopatologia de La Inflamacióncaritoandrea16100% (1)
- Sistema inmunológico: definición, tipos de defensa, antígeno y anticuerposDocumento12 páginasSistema inmunológico: definición, tipos de defensa, antígeno y anticuerposJuan Ignacio D'AversaAún no hay calificaciones
- InflamaciónDocumento18 páginasInflamaciónOscar Argüello100% (1)
- Inmunidad InnataDocumento23 páginasInmunidad InnataMarian SotoAún no hay calificaciones
- Inmunología humana: Sistema inmune innato y adaptativoDocumento96 páginasInmunología humana: Sistema inmune innato y adaptativoKadh MenezesAún no hay calificaciones
- Inflamación aguda: causas, mecanismos y evoluciónDocumento9 páginasInflamación aguda: causas, mecanismos y evoluciónJosé ManuelAún no hay calificaciones
- Patologia de La InflamaciónDocumento57 páginasPatologia de La InflamaciónLizeth GarciaAún no hay calificaciones
- fisiologia resistencia a la infecciónDocumento11 páginasfisiologia resistencia a la infecciónDani Slayer IriarteAún no hay calificaciones
- Resumen de Inflamación Patologia Cap2 Robbins 8va EdDocumento8 páginasResumen de Inflamación Patologia Cap2 Robbins 8va EdjordanodmAún no hay calificaciones
- Taller 1 Patología 2021-IDocumento4 páginasTaller 1 Patología 2021-ICarmen Claudia Buitrago PorrasAún no hay calificaciones
- Fisioterapia InflamacionDocumento17 páginasFisioterapia InflamacionCristobal Hernandez MadridAún no hay calificaciones
- InmudidadDocumento34 páginasInmudidadFabrizzio RomeroAún no hay calificaciones
- Expo Patologia FinalDocumento5 páginasExpo Patologia FinalAlexandra castroAún no hay calificaciones
- Sistema inmunitario: definición, componentes y mecanismos de defensaDocumento24 páginasSistema inmunitario: definición, componentes y mecanismos de defensaYolandaAún no hay calificaciones
- InflamaciónDocumento146 páginasInflamaciónDane RuedaAún no hay calificaciones
- Guia InmunologiaDocumento15 páginasGuia InmunologiaYeraAún no hay calificaciones
- INFLAMACIÓNDocumento8 páginasINFLAMACIÓNDavid TrejosAún no hay calificaciones
- Inflamación AgudaDocumento4 páginasInflamación AgudaNathaly BaldiosedaAún no hay calificaciones
- RESUMEN Inmunidad antibacterianaDocumento6 páginasRESUMEN Inmunidad antibacterianaLetycia GodoyAún no hay calificaciones
- Lectura Proceso - InflamatorioDocumento7 páginasLectura Proceso - Inflamatorioeudicsalazar2Aún no hay calificaciones
- Inflamación aguda y crónicaDocumento18 páginasInflamación aguda y crónicaYanina Pdv90% (10)
- Inflamación Aguda y CrónicaDocumento6 páginasInflamación Aguda y CrónicaFernando Villavicencio100% (1)
- Sistema LinfáticoDocumento11 páginasSistema LinfáticoJuan de diosAún no hay calificaciones
- Eje 1 Sistema Inmune Mecanismos InespecificosDocumento7 páginasEje 1 Sistema Inmune Mecanismos InespecificosMary Tamara JimenezAún no hay calificaciones
- Agabriela - Arriaga - Unidada 2 - Actividad 2 - Inflamacion Aguda y CronicaDocumento12 páginasAgabriela - Arriaga - Unidada 2 - Actividad 2 - Inflamacion Aguda y Cronicadaniel chaparroAún no hay calificaciones
- Mecanismo de Respuesta Inmune Del HuéspedDocumento7 páginasMecanismo de Respuesta Inmune Del Huésped99twtqjsxyAún no hay calificaciones
- Inflamación AgudaDocumento4 páginasInflamación AgudaOriana RegaladoAún no hay calificaciones
- Inflamacion InmunologiaDocumento33 páginasInflamacion InmunologiaDiana Arely Cervantes GonzálezAún no hay calificaciones
- Resumen Inflamacion Patologia RobbinsDocumento8 páginasResumen Inflamacion Patologia RobbinsDennisse Pinto100% (4)
- Fases de La InflamacionDocumento8 páginasFases de La InflamacionandresAún no hay calificaciones
- Resumen de InmunidadDocumento10 páginasResumen de InmunidadLisbeth Crespo CuencaAún no hay calificaciones
- Atlas de InmunologiaDocumento50 páginasAtlas de InmunologiaNuñez Fonseca Jose100% (1)
- Acontecimientos CelularesDocumento7 páginasAcontecimientos CelularesNico Blanco BarajasAún no hay calificaciones
- 001 Sris y SdmoDocumento6 páginas001 Sris y Sdmovitorinno5705100% (2)
- Taller Preparatorio Primer Parcial 2 UscDocumento21 páginasTaller Preparatorio Primer Parcial 2 UscTatiana Guzmán SalinasAún no hay calificaciones
- Síndrome de Respuesta Inflamatoria SistémicaDocumento6 páginasSíndrome de Respuesta Inflamatoria Sistémicalucero166Aún no hay calificaciones
- QUimiocinasDocumento5 páginasQUimiocinasJose Renato GonzálezAún no hay calificaciones
- Inflamación: Mecanismos e inmunologíaDocumento26 páginasInflamación: Mecanismos e inmunologíaAbner Hernandez LopezAún no hay calificaciones
- Diarrea Aguda AEPDocumento10 páginasDiarrea Aguda AEPcpfredAún no hay calificaciones
- Clinica DengueDocumento46 páginasClinica DengueJuan Carlos FernandezAún no hay calificaciones
- Evolucion Historica Del Concepto y Criterios ActuaDocumento15 páginasEvolucion Historica Del Concepto y Criterios ActuaRaed OrtizAún no hay calificaciones
- Clasificacion, Diagnostico y Tratamiento Integral Del DengueDocumento13 páginasClasificacion, Diagnostico y Tratamiento Integral Del DengueCarlosLópezAAún no hay calificaciones
- Alteraciones Circadianas Del Sistema CardiovascularDocumento6 páginasAlteraciones Circadianas Del Sistema CardiovascularRaed OrtizAún no hay calificaciones
- MeningitisBacterianaSintomasDiagnosticoDocumento11 páginasMeningitisBacterianaSintomasDiagnosticoEmily MariñosAún no hay calificaciones
- Fisiopatologia de La Inflamacion FarmacoDocumento17 páginasFisiopatologia de La Inflamacion FarmacoRaed OrtizAún no hay calificaciones
- Norma 147 - Manejo Del DengueDocumento111 páginasNorma 147 - Manejo Del Denguemcookie250% (2)
- Tesis ListoDocumento56 páginasTesis ListoRaed OrtizAún no hay calificaciones
- RMQ 052 JDocumento5 páginasRMQ 052 JRaed OrtizAún no hay calificaciones
- Instrucciones LCRDocumento1 páginaInstrucciones LCRRaed OrtizAún no hay calificaciones
- Dolor AbdominalDocumento10 páginasDolor AbdominalOmarAún no hay calificaciones
- Neumotorax, PneumotoraxDocumento10 páginasNeumotorax, PneumotoraxJorge VergaraAún no hay calificaciones
- HELICOBACTER PYLORI - Clinica, Diagnostico y TratamientoDocumento4 páginasHELICOBACTER PYLORI - Clinica, Diagnostico y TratamientomasterchipAún no hay calificaciones
- Poli Trauma AbdominalDocumento12 páginasPoli Trauma AbdominalRaed OrtizAún no hay calificaciones
- Secreción Biliar y Mecanismos de Digestión de Grasas 2017-I PDFDocumento26 páginasSecreción Biliar y Mecanismos de Digestión de Grasas 2017-I PDFLuz BecerraAún no hay calificaciones
- Instrucciones LCRDocumento1 páginaInstrucciones LCRRaed OrtizAún no hay calificaciones
- Shock neurogénico: fisiopatología, diagnóstico y tratamientoDocumento5 páginasShock neurogénico: fisiopatología, diagnóstico y tratamientoChristian MolinaAún no hay calificaciones
- Hi Peru Rice MiaDocumento1 páginaHi Peru Rice MiaRocío García OgallaAún no hay calificaciones
- SIKADocumento25 páginasSIKANahin ChumaceroAún no hay calificaciones
- Escala Child PughDocumento2 páginasEscala Child PughAntonio ZaldivarAún no hay calificaciones
- Malaria Treatment Guidelines 2011 Esp PDFDocumento163 páginasMalaria Treatment Guidelines 2011 Esp PDFRaed OrtizAún no hay calificaciones
- Revchilanestv40n04 04 PDFDocumento19 páginasRevchilanestv40n04 04 PDFRaed OrtizAún no hay calificaciones
- Alteraciones Circadianas Del Sistema CardiovascularDocumento6 páginasAlteraciones Circadianas Del Sistema CardiovascularRaed OrtizAún no hay calificaciones
- Intoxicaciones No FarmacologicasDocumento13 páginasIntoxicaciones No FarmacologicasArianna Rodríguez ContrerasAún no hay calificaciones
- Tema 11-Bloque II-La Piel. Estructura y FuncionesDocumento7 páginasTema 11-Bloque II-La Piel. Estructura y FuncionesGenesis Gonzalez ReyesAún no hay calificaciones
- 1 Farmacologia Betabloqueantes PDFDocumento16 páginas1 Farmacologia Betabloqueantes PDFIzadora CorrêaAún no hay calificaciones
- Estudio Etiológico y Manejo de La Anafilaxia - MedwaveDocumento14 páginasEstudio Etiológico y Manejo de La Anafilaxia - MedwaveRaed OrtizAún no hay calificaciones
- Rer Sepsis Puerperal PDFDocumento62 páginasRer Sepsis Puerperal PDFFer Rebolledo AvilésAún no hay calificaciones
- MetrorragiasDocumento3 páginasMetrorragiasMilton Gary Tapia VargasAún no hay calificaciones
- Mutacion Del AdnDocumento9 páginasMutacion Del AdnKevin J CevallosAún no hay calificaciones
- Biologia II RemovedDocumento18 páginasBiologia II RemovedMonserrat LizarragaAún no hay calificaciones
- Corolario de ParasitologíaDocumento2 páginasCorolario de ParasitologíaAnonymous oHZRC9R1Aún no hay calificaciones
- Informe MorfologiaDocumento12 páginasInforme Morfologia『カルロス・レオン』Aún no hay calificaciones
- Ecosistema TripticoDocumento2 páginasEcosistema TripticoSisley Filos60% (5)
- Practica N 7Documento4 páginasPractica N 7Jonathan Andres MolanoAún no hay calificaciones
- LisosomasDocumento2 páginasLisosomasEdwin TaramuelAún no hay calificaciones
- Endocrinología Del EmbarazoDocumento27 páginasEndocrinología Del EmbarazoKeila CalixtoAún no hay calificaciones
- Célula VegetalDocumento27 páginasCélula VegetalDaniela MeloAún no hay calificaciones
- AGUAYMANTODocumento14 páginasAGUAYMANTOMayra Ih Jiménez PastorAún no hay calificaciones
- Guía 3 de Aprendizaje Virtua-2021 Biología Grado OctavoDocumento4 páginasGuía 3 de Aprendizaje Virtua-2021 Biología Grado Octavojose valderramaAún no hay calificaciones
- Función y estructura del sistema nervioso centralDocumento5 páginasFunción y estructura del sistema nervioso centralEfrain CuellarAún no hay calificaciones
- Temario Biologia IPN Area MatDocumento3 páginasTemario Biologia IPN Area MatJuan RodriguezAún no hay calificaciones
- Fisiología Del Músculo EsqueléticoDocumento18 páginasFisiología Del Músculo EsqueléticoVICTOR HUGO OLORTEGUI GARCIAAún no hay calificaciones
- Ejercicios PDFDocumento14 páginasEjercicios PDFJose Miguel Cotes TorresAún no hay calificaciones
- Apendicitis AgudaDocumento10 páginasApendicitis AgudaCatha LozadaAún no hay calificaciones
- Síndrome de KlinefelterDocumento7 páginasSíndrome de Klinefeltermoises orozcoAún no hay calificaciones
- Rabia Tabla 2021Documento23 páginasRabia Tabla 2021CesarFloresAún no hay calificaciones
- PATRONES DE COLORDocumento1 páginaPATRONES DE COLORSofia JaimesAún no hay calificaciones
- Estudios de Bosques en La Region CajamarcaDocumento18 páginasEstudios de Bosques en La Region CajamarcaKellisasha LozanoAún no hay calificaciones
- Repaso Bio - Parte IDocumento34 páginasRepaso Bio - Parte IPaulana OregarAún no hay calificaciones
- RespuestaConsolidación Orientadora 5Documento3 páginasRespuestaConsolidación Orientadora 5Maxis GonzalezAún no hay calificaciones
- AaP C07 U1 PlaneadorDocumento15 páginasAaP C07 U1 PlaneadorcandyAún no hay calificaciones
- Evaluacion BioenergeticaDocumento3 páginasEvaluacion BioenergeticaEstefania De Battista MoroAún no hay calificaciones
- Respiración celular: proceso de obtención de ATPDocumento1 páginaRespiración celular: proceso de obtención de ATPANDREA MICHELLE PORTILLO PINEDA100% (1)
- Árboles filogenéticos, grupos monofiléticos y clasificación de especiesDocumento2 páginasÁrboles filogenéticos, grupos monofiléticos y clasificación de especiesGEANPIERRE ENRIQUE MAURICIO CASTROAún no hay calificaciones
- El Mapa Conceptual y La InfografíaDocumento20 páginasEl Mapa Conceptual y La InfografíaangieAún no hay calificaciones
- Patologia GeneralDocumento102 páginasPatologia GeneralBastianAún no hay calificaciones
- GatosDocumento3 páginasGatosMomoko TejidosAún no hay calificaciones
- Métodos de Medición de Crecimiento CelularDocumento9 páginasMétodos de Medición de Crecimiento CelularYesenia FelixAún no hay calificaciones