También podría gustarte
- Secuencias Segunda Etapa Del AñoDocumento35 páginasSecuencias Segunda Etapa Del AñoÁlvaro ZapataAún no hay calificaciones
- Resumen Membranas BiologicasDocumento6 páginasResumen Membranas BiologicaslizzyrodnuzAún no hay calificaciones
- 01 Membrana CelularDocumento22 páginas01 Membrana CelularMarco ChavezAún no hay calificaciones
- Introduccion - Tipos de MembranaDocumento9 páginasIntroduccion - Tipos de MembranaJose Luis CTAún no hay calificaciones
- Temas 7 y 8 Estructura de La Célula EucarióticaDocumento12 páginasTemas 7 y 8 Estructura de La Célula EucarióticaLuis Hernández PeñaAún no hay calificaciones
- TEMA 7. Envolturas de La CélulaDocumento7 páginasTEMA 7. Envolturas de La CélulaSara AltunaAún no hay calificaciones
- Membrana Plasmatica e Interacciones CelularesDocumento14 páginasMembrana Plasmatica e Interacciones CelularesKatty RodriguezAún no hay calificaciones
- Tema 4. La Estructura de La CélulaDocumento6 páginasTema 4. La Estructura de La CélulaPablo Martos AguayoAún no hay calificaciones
- Membrana CelularDocumento32 páginasMembrana Celularisi valdiviaAún no hay calificaciones
- Nivel CelularDocumento18 páginasNivel CelularJoaquinLuiniAún no hay calificaciones
- Tema 27 Precege PruebaDocumento9 páginasTema 27 Precege PruebaIre del CerroAún no hay calificaciones
- Celula Patologia Clinica 23-09-2023Documento25 páginasCelula Patologia Clinica 23-09-2023JUANA ADELA HUANCA CALLISAYAAún no hay calificaciones
- Celula Patologia Clinica 24-09-2023Documento25 páginasCelula Patologia Clinica 24-09-2023JUANA ADELA HUANCA CALLISAYAAún no hay calificaciones
- Tema 8: "Reproducción Celular.": Tema 9: "Membrana Plasmática Y Orgánulos Membranosos."Documento24 páginasTema 8: "Reproducción Celular.": Tema 9: "Membrana Plasmática Y Orgánulos Membranosos."mariaAún no hay calificaciones
- BLOQUE I. La CélulaDocumento9 páginasBLOQUE I. La Célulanoedrin2Aún no hay calificaciones
- Membrana CelularDocumento22 páginasMembrana CelularManuel Cespedes100% (1)
- Los Orgánulos CelularesDocumento15 páginasLos Orgánulos CelularesamandaAún no hay calificaciones
- 2º Bachillerato, Tema 6 (Membrana y Envolturas Celulares)Documento9 páginas2º Bachillerato, Tema 6 (Membrana y Envolturas Celulares)ms marvelAún no hay calificaciones
- Organización de La Celula KeikoDocumento10 páginasOrganización de La Celula KeikoClaribel Yovera CarrilloAún no hay calificaciones
- 06-Organización Celular I-SEDocumento15 páginas06-Organización Celular I-SEhyrnetsvvAún no hay calificaciones
- Esquema Resumen 7Documento15 páginasEsquema Resumen 7Jennifer Diaz PomaAún no hay calificaciones
- 8.membrana, PC y CitopDocumento72 páginas8.membrana, PC y CitopEmma Cruz López de PabloAún no hay calificaciones
- Guia La Celula-Membrana Celular 2017 PDFDocumento15 páginasGuia La Celula-Membrana Celular 2017 PDFjusto pastor sarmiento cstañedaAún no hay calificaciones
- FISIOLOGIADocumento5 páginasFISIOLOGIARosivell RivasAún no hay calificaciones
- Bio b2 - Removed PDFDocumento25 páginasBio b2 - Removed PDFRamon Garcia DíazAún no hay calificaciones
- 04 Membrana PlasmáticaDocumento14 páginas04 Membrana PlasmáticaCamila RamirezAún no hay calificaciones
- Guia 2Documento13 páginasGuia 2pabloAún no hay calificaciones
- Biologia General Membrana y TransportesDocumento5 páginasBiologia General Membrana y TransportesBlankyz LunaAún no hay calificaciones
- Práctica 1° Medio InternoDocumento22 páginasPráctica 1° Medio InternoMiguel Andrex Albix Florx0% (1)
- En Volturas Celulares 2º BachDocumento5 páginasEn Volturas Celulares 2º BachMónica Fernández SantosAún no hay calificaciones
- La Membrana CelularDocumento21 páginasLa Membrana CelularPablo Cesar Gutierrez Del VillarAún no hay calificaciones
- Resumen Final de Zoología GeneralDocumento47 páginasResumen Final de Zoología Generalisabellacb19Aún no hay calificaciones
- G C8 Membrana BCM SA2023Documento55 páginasG C8 Membrana BCM SA2023Chiluisa EdisonAún no hay calificaciones
- Módulo de Biología de 10 #2Documento6 páginasMódulo de Biología de 10 #2HAFFID BALLESTEROSAún no hay calificaciones
- T 6. La Célula y Las Envolturas CelularesDocumento11 páginasT 6. La Célula y Las Envolturas CelularesIrene García MarcosAún no hay calificaciones
- 08 Envolturas CelularesDocumento8 páginas08 Envolturas CelularesSusana FloAún no hay calificaciones
- Membrana Plasmatica 1Documento40 páginasMembrana Plasmatica 1areli elizabeth reynoso jimenezAún no hay calificaciones
- Unidad 2 CitoplasmaDocumento34 páginasUnidad 2 CitoplasmaAndrea DuránAún no hay calificaciones
- Modulo 2Documento6 páginasModulo 2diegoAún no hay calificaciones
- Modulo 2Documento6 páginasModulo 2diego100% (1)
- Estructura y Función en Procariotas y EucariotasDocumento87 páginasEstructura y Función en Procariotas y EucariotasempresvarAún no hay calificaciones
- Tema 7. La Membrana CelularDocumento7 páginasTema 7. La Membrana CelularVictor AlonsoAún no hay calificaciones
- Universidad Nacional Abierta y A Distancia Vicerrectoría Académica y de Investigación Curso SolucionDocumento4 páginasUniversidad Nacional Abierta y A Distancia Vicerrectoría Académica y de Investigación Curso SolucionJose ValenciaAún no hay calificaciones
- Membrana PlasmáticaDocumento7 páginasMembrana PlasmáticaFerni MoralesAún no hay calificaciones
- Objetivo 3. Transporte CelularDocumento7 páginasObjetivo 3. Transporte Celularmusicart30008394100% (2)
- 3 - Célula y Membrana CelularDocumento32 páginas3 - Célula y Membrana Celularjose.torres1Aún no hay calificaciones
- CONT. TEMA 4 BIOL. 5to. BDocumento4 páginasCONT. TEMA 4 BIOL. 5to. BDeynor CopaAún no hay calificaciones
- Biolo 03Documento8 páginasBiolo 03Jorge RopónAún no hay calificaciones
- La Membrana CelularDocumento22 páginasLa Membrana CelularJorge AndrésAún no hay calificaciones
- Test de BiologiaDocumento11 páginasTest de BiologiaCamila G. GutiérrezAún no hay calificaciones
- Transporte de MembranaDocumento5 páginasTransporte de MembranaPaula LopezAún no hay calificaciones
- Membrana Celular Mapa Conceptual..... Norma 5..Documento8 páginasMembrana Celular Mapa Conceptual..... Norma 5..Jamirton Erledy GarciaAún no hay calificaciones
- Material 2019D1 BIO101 01 115727Documento30 páginasMaterial 2019D1 BIO101 01 115727Jaime HuidobroAún no hay calificaciones
- Biología Tema 2Documento8 páginasBiología Tema 2emilu escobar romeroAún no hay calificaciones
- Lectura Semana 1 - La CelulaDocumento10 páginasLectura Semana 1 - La CelulashakaAún no hay calificaciones
- TALLER BIOLOGÍA ImprimirDocumento5 páginasTALLER BIOLOGÍA ImprimirRocketcat100% (4)
- T 1 Biología La CélulaDocumento63 páginasT 1 Biología La CélulaAlondra Cabezudo JuarezAún no hay calificaciones
- Taller 4 - La Membrana CelularDocumento8 páginasTaller 4 - La Membrana CelularJuan Carlos Portillo VillarruelAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Tejidos Conectivos de Consistencia Rígida 1Documento18 páginasTejidos Conectivos de Consistencia Rígida 1isabella machoAún no hay calificaciones
- Filamentos IntermediosDocumento8 páginasFilamentos Intermediosisabella machoAún no hay calificaciones
- Tejido EpitelialDocumento6 páginasTejido Epitelialisabella machoAún no hay calificaciones
- Carta de SolteriaDocumento1 páginaCarta de Solteriaisabella macho100% (2)
- Documento de Transporte 1300000009 XLK332 PDFDocumento1 páginaDocumento de Transporte 1300000009 XLK332 PDFJudisita Ce AAún no hay calificaciones
- Trabajo de Unidad 1 - Sistema de Ecuaciones LinealesDocumento10 páginasTrabajo de Unidad 1 - Sistema de Ecuaciones LinealesPaula Sirlopu FachoAún no hay calificaciones
- Nuevo Preparador de Jardin 3 PeriodoDocumento28 páginasNuevo Preparador de Jardin 3 Periodoluz marina jimenez martinezAún no hay calificaciones
- E P RDocumento13 páginasE P RWilliam Anthony Chapoñan CajusolAún no hay calificaciones
- Cartilla de Junio 2do C de TecnologiaDocumento12 páginasCartilla de Junio 2do C de Tecnologiamarina barriosAún no hay calificaciones
- Homilía Lunes Santo 6 de Abril de 2020Documento2 páginasHomilía Lunes Santo 6 de Abril de 2020Alexander Alvarado MalaverAún no hay calificaciones
- Solución Práctica 4 Bancos (Contabilidad Intermedia)Documento6 páginasSolución Práctica 4 Bancos (Contabilidad Intermedia)Jhoselin Mamani Ticona100% (2)
- Analisis de SassouniDocumento59 páginasAnalisis de SassouniPaola Andrea Quintero BahamonAún no hay calificaciones
- Cantidad de Nieve Promedio en El Estado Colorado InglesDocumento2 páginasCantidad de Nieve Promedio en El Estado Colorado InglesSebastian HerreraAún no hay calificaciones
- Mapa Conceptual-Arte Imagen y Saber AncestralDocumento3 páginasMapa Conceptual-Arte Imagen y Saber AncestralDaniela Blanco SerpaAún no hay calificaciones
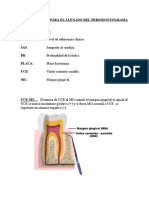
- EXPLICACION Periodontograma IDocumento4 páginasEXPLICACION Periodontograma IYochito75% (4)
- Programacion Flota Nacional 17-04-2019Documento54 páginasProgramacion Flota Nacional 17-04-2019Claudio Felipe Hernández BrunaAún no hay calificaciones
- Sistemas de Motores DieselDocumento34 páginasSistemas de Motores DieselJORGE ANTONIO MAY CRUZAún no hay calificaciones
- Evaluacion Tecnica de Raise BoringDocumento8 páginasEvaluacion Tecnica de Raise BoringFelimon William Espinoza Huaman0% (1)
- Práctica Calificada Respiracion ExcrecionDocumento12 páginasPráctica Calificada Respiracion ExcrecionQuintín Esteban PacoAún no hay calificaciones
- PENCICLOVIRDocumento27 páginasPENCICLOVIRDiego Llusco CalleAún no hay calificaciones
- Manual MultisimDocumento43 páginasManual MultisimJhonny Emilio CervantesAún no hay calificaciones
- Salud Publica Examen 2 ParcialDocumento13 páginasSalud Publica Examen 2 ParcialEduardo Bonifacio MoralesAún no hay calificaciones
- Autoestima 2Documento30 páginasAutoestima 2Yemina Rosalía Dupertuis BalpredaAún no hay calificaciones
- Nelcy Robles 1Documento124 páginasNelcy Robles 1Carlos AncasiAún no hay calificaciones
- La Sabiduría de La CruzDocumento4 páginasLa Sabiduría de La CruzdanielpablokmAún no hay calificaciones
- Alteraciones Agudas de Las Transaminasas y ColestasisDocumento13 páginasAlteraciones Agudas de Las Transaminasas y ColestasisAdrian CrespoAún no hay calificaciones
- Medicina Estética M2T7Documento8 páginasMedicina Estética M2T7CHRISTIANAún no hay calificaciones
- MonIRQ4. Baterías CSTRDocumento2 páginasMonIRQ4. Baterías CSTRAaron CarvajalAún no hay calificaciones
- Resumen 1 Enfermedades RespiratoriasDocumento24 páginasResumen 1 Enfermedades RespiratoriasJaime Andrés Bravo Tapia100% (1)
- Creacion de Un Cluster Virtual Con ABC LinuxDocumento5 páginasCreacion de Un Cluster Virtual Con ABC LinuxKelvin VelezAún no hay calificaciones
- Anexo ADocumento4 páginasAnexo AMackarena QuirogaAún no hay calificaciones
- Acelerador Magnético (Rifle de Gauss) PDFDocumento2 páginasAcelerador Magnético (Rifle de Gauss) PDFChrisXYZ147100% (1)
- Levantamiento Observaciones EstructurasDocumento8 páginasLevantamiento Observaciones EstructurasJosef Walter SosaAún no hay calificaciones
- Adultez JovenDocumento6 páginasAdultez JovenCarlig LeonAún no hay calificaciones