También podría gustarte
- Introducción a la química de los metales de transiciónDe EverandIntroducción a la química de los metales de transiciónAún no hay calificaciones
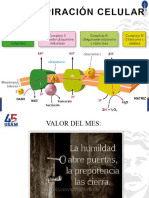
- Cadena Respiratoria (Texto Estudio) y Fosforilacion OxidativaDocumento13 páginasCadena Respiratoria (Texto Estudio) y Fosforilacion Oxidativajdesiree1803Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
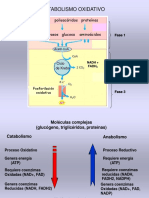
- Fosforilación OxidativaDocumento2 páginasFosforilación OxidativaNEIVERAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- TEORICO Cadena Resp, Fosfor OxidativaDocumento12 páginasTEORICO Cadena Resp, Fosfor OxidativaSofia CoriaAún no hay calificaciones
- Acondicionado de materiales termoplásticos para su transformación. QUIT0209De EverandAcondicionado de materiales termoplásticos para su transformación. QUIT0209Aún no hay calificaciones
- 3-2022 Cte y Fosforilacion OxidativaDocumento56 páginas3-2022 Cte y Fosforilacion OxidativaAbisai FuentesAún no hay calificaciones
- Conferencia 17 La Fosforilación OxidativaDocumento33 páginasConferencia 17 La Fosforilación OxidativaJoaquim JosephAún no hay calificaciones
- Fosforilacion OxidativaDocumento6 páginasFosforilacion Oxidativalore05Aún no hay calificaciones
- Clase 6 Fosforilación Oxidativa y Fotofosforilación (TEM4018)Documento18 páginasClase 6 Fosforilación Oxidativa y Fotofosforilación (TEM4018)constvnza16Aún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento14 páginasCadena Respiratoria y Fosforilación OxidativaP. Arevalo IlianaAún no hay calificaciones
- 003 Bioenergética UC 2020Documento30 páginas003 Bioenergética UC 2020Helena QuintanaAún no hay calificaciones
- Discu 4Documento4 páginasDiscu 4alaya vasquezAún no hay calificaciones
- Cadena Respiratoria Ciclo 1-2020Documento39 páginasCadena Respiratoria Ciclo 1-2020Grace AlvaradoAún no hay calificaciones
- Resolución Del Taller 5 - Cadena Respiratoria y Fosforilación OxidativaDocumento15 páginasResolución Del Taller 5 - Cadena Respiratoria y Fosforilación Oxidativassmrt.2016Aún no hay calificaciones
- Practica 07. Inhibidores de Procesos BioenergeticosDocumento5 páginasPractica 07. Inhibidores de Procesos BioenergeticosEsthyv Loyaga PonceAún no hay calificaciones
- Cadena Respiratoria Fosforilacion OxidatDocumento95 páginasCadena Respiratoria Fosforilacion OxidatMiguel Soni CasillasAún no hay calificaciones
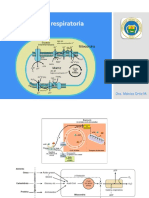
- Transporte de ELectrones y Fosforilacion OxidativaDocumento16 páginasTransporte de ELectrones y Fosforilacion OxidativaEte64Aún no hay calificaciones
- Esquema de Cadena Transportadora de El Ectrones y Fosforilación OxidativaDocumento4 páginasEsquema de Cadena Transportadora de El Ectrones y Fosforilación OxidativaJuanPérezAún no hay calificaciones
- CTE y FO para BlogDocumento9 páginasCTE y FO para BlogcalpainaAún no hay calificaciones
- Bioenergetica Fotosintesis 2019Documento51 páginasBioenergetica Fotosintesis 2019Antonela SorucoAún no hay calificaciones
- Cuestionario de Bioenergética, Metabolismo y AguaDocumento18 páginasCuestionario de Bioenergética, Metabolismo y Agualuis javierAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento5 páginasCadena Transportadora de ElectronesVania MarínAún no hay calificaciones
- Biología I Tema 3.2.5.3 Cadena RespiratoriaDocumento30 páginasBiología I Tema 3.2.5.3 Cadena RespiratoriaTita Oz100% (1)
- Apuntes de Cadenas Respiratorias.Documento6 páginasApuntes de Cadenas Respiratorias.Proto 2019Aún no hay calificaciones
- Cadena Respiratoria, Fosforilacion Oxidativa Cap. 8 FerrierDocumento5 páginasCadena Respiratoria, Fosforilacion Oxidativa Cap. 8 FerrierEvelyn Guadalupe González BravoAún no hay calificaciones
- Fosforilación OxidativaDocumento20 páginasFosforilación OxidativaValeri Vasquez Cucho0% (1)
- Transporte Electronico y Fosforilacion OxidativaDocumento41 páginasTransporte Electronico y Fosforilacion OxidativanicolehurtadosAún no hay calificaciones
- Ba Res 13Documento6 páginasBa Res 13Rudan Retamozo OrihuelaAún no hay calificaciones
- Ciclo de Krebs y Cadena RespDocumento7 páginasCiclo de Krebs y Cadena RespLuciana AtencioAún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento2 páginasCadena Respiratoria y Fosforilación OxidativaDarikson FaustoAún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento6 páginasCadena Respiratoria y Fosforilación OxidativahenryAún no hay calificaciones
- 06-Fosforilación OxidativaDocumento4 páginas06-Fosforilación OxidativaYasmin Morales LópezAún no hay calificaciones
- Cadena RespiratoriaDocumento25 páginasCadena RespiratoriaKruzkaya MachicadoAún no hay calificaciones
- Fosforilación OxidativaDocumento3 páginasFosforilación Oxidativafrancisco valdebenitoAún no hay calificaciones
- Principales Rutas CatabólicasDocumento49 páginasPrincipales Rutas CatabólicasReben ZecnajAún no hay calificaciones
- Fosforilacion OxidativaDocumento25 páginasFosforilacion OxidativaEvelyn Guadalupe BravoAún no hay calificaciones
- Fosforilacion OxidativaDocumento5 páginasFosforilacion Oxidativaaoihana18100% (1)
- Bioenergetica y Fosforilacion OxidativaDocumento4 páginasBioenergetica y Fosforilacion OxidativaFernanda MejiaAún no hay calificaciones
- Cadena Trasportadora D ElectronesDocumento45 páginasCadena Trasportadora D ElectronesKendy SahibAún no hay calificaciones
- Seminario Fosforilación OxidativaDocumento3 páginasSeminario Fosforilación Oxidativabelirosi90Aún no hay calificaciones
- Resumen Completo de La Cadena Transportadora de Electrones y Fosforilación OxidativaDocumento3 páginasResumen Completo de La Cadena Transportadora de Electrones y Fosforilación OxidativaCarolina DuranAún no hay calificaciones
- Semana 7Documento47 páginasSemana 7Marilyn TantaquispeAún no hay calificaciones
- Fase Dependiente de La Luz o FotoquímicaDocumento3 páginasFase Dependiente de La Luz o FotoquímicaJosue RebolledoAún no hay calificaciones
- Cadena RespiratoriaDocumento26 páginasCadena Respiratoriales12michuAún no hay calificaciones
- Fosforilación Oxidativa Ciclo II-2023Documento22 páginasFosforilación Oxidativa Ciclo II-2023Ericson CastroAún no hay calificaciones
- Cadena de Trasporte y Fosforilacion KKDocumento32 páginasCadena de Trasporte y Fosforilacion KKUriel AlvaradoAún no hay calificaciones
- TRANSPORTE DE ELECTRONES Y FOSFORILACION OXIDATIVAsegundoparcialpdfDocumento39 páginasTRANSPORTE DE ELECTRONES Y FOSFORILACION OXIDATIVAsegundoparcialpdfDebora RuizAún no hay calificaciones
- Fosforilacion OxidativaDocumento16 páginasFosforilacion OxidativaFlorencia BarsiAún no hay calificaciones
- Cadena Transporte de ElectronesDocumento14 páginasCadena Transporte de ElectronesAleKcis GarCiaAún no hay calificaciones
- U4 Act 2. Cadena de Transporte de Electrones - 2Documento6 páginasU4 Act 2. Cadena de Transporte de Electrones - 2Rocío P AscAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento5 páginasCadena Transportadora de ElectronesAmadaGarciaYepezAún no hay calificaciones
- Clase 9.2Documento28 páginasClase 9.2Guadalupe CarhuatantaAún no hay calificaciones
- BioquímicaDocumento12 páginasBioquímicaJenniferAún no hay calificaciones
- Teoria QuimiosDocumento11 páginasTeoria QuimiosJoaquin SampedroAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento16 páginasCadena Transportadora de ElectronesELIZABETH SMAún no hay calificaciones
- Transporte Electrónico y Fosforilación OxidativaDocumento33 páginasTransporte Electrónico y Fosforilación OxidativaJimmy ArroligaAún no hay calificaciones
- La Fosforilación OxidativaDocumento8 páginasLa Fosforilación OxidativaJose JuanAún no hay calificaciones
- Respiración Celular y Transferencia de Energía BiológicaDocumento27 páginasRespiración Celular y Transferencia de Energía BiológicakcaceresmaAún no hay calificaciones
- RESUMEN OBSTETRICIA MF A CompletarDocumento69 páginasRESUMEN OBSTETRICIA MF A Completardocsamu78Aún no hay calificaciones
- Cuestionario Redox v0.4 PDFDocumento7 páginasCuestionario Redox v0.4 PDFdocsamu78Aún no hay calificaciones
- Patología CelularDocumento44 páginasPatología Celulardocsamu78Aún no hay calificaciones
- Ciclo de Krebs y LanzaderasDocumento52 páginasCiclo de Krebs y Lanzaderasdocsamu78100% (1)
- 3 Er Parcial Bioquimica 2017 CompletoDocumento267 páginas3 Er Parcial Bioquimica 2017 Completodocsamu780% (1)
- Bioest JuriDocumento95 páginasBioest Juridocsamu78Aún no hay calificaciones
- TP 12 - Cetogénesis y Cetolisis. Cetoacidosis - 2015Documento6 páginasTP 12 - Cetogénesis y Cetolisis. Cetoacidosis - 2015docsamu78Aún no hay calificaciones
- TP 11 - Síntesis y Degradación de Triglicéridos - 2015Documento7 páginasTP 11 - Síntesis y Degradación de Triglicéridos - 2015docsamu780% (1)
- Bbiq U1 Ea Maas.Documento5 páginasBbiq U1 Ea Maas.Martha Maria Araiza SantiniAún no hay calificaciones
- Manual de Fisio y BioqDocumento82 páginasManual de Fisio y BioqYozzalet LGAún no hay calificaciones
- Guia 5Documento15 páginasGuia 5Lucas Ezequiel AstudilloAún no hay calificaciones
- Fosforilacion Oxidativa y Cadena RespiratoriaDocumento19 páginasFosforilacion Oxidativa y Cadena RespiratoriaerikaAún no hay calificaciones
- Cadena Respiratoria Fosforilacion OxidatDocumento95 páginasCadena Respiratoria Fosforilacion OxidatMiguel Soni CasillasAún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento6 páginasCadena Respiratoria y Fosforilación OxidativahenryAún no hay calificaciones
- 5 Mitocondria y EnergiaDocumento11 páginas5 Mitocondria y EnergiaGaston MoralesAún no hay calificaciones
- Actividad 3. - Diagrama de Flujo de La FotosintesisDocumento4 páginasActividad 3. - Diagrama de Flujo de La Fotosintesiszorayadallana82% (11)
- ATP AsaDocumento6 páginasATP AsaMiguel LópezAún no hay calificaciones
- CLASE 4 - Defi G6PD - DM - Cad Respiratoria DiabetesDocumento38 páginasCLASE 4 - Defi G6PD - DM - Cad Respiratoria DiabetesOrions MTovalinoAún no hay calificaciones
- Resumen Robertis 2do ParcialDocumento15 páginasResumen Robertis 2do ParcialYohana SalazarAún no hay calificaciones
- Tp04 de Bioquímica UNLPDocumento7 páginasTp04 de Bioquímica UNLPAlyson GuilhermeAún no hay calificaciones
- Cuestionario de Bioenergética, Metabolismo y AguaDocumento18 páginasCuestionario de Bioenergética, Metabolismo y Agualuis javierAún no hay calificaciones
- Mitocondrias y Cloroplastos If 4Documento21 páginasMitocondrias y Cloroplastos If 4CHRISTIAN ANDRES MORENO CHIMBORAún no hay calificaciones
- BIOQUIMICADocumento113 páginasBIOQUIMICACarolina CAún no hay calificaciones
- Fosforilación OxidativaDocumento25 páginasFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezAún no hay calificaciones
- Qué Es Una Proteína Desacopladora PDFDocumento5 páginasQué Es Una Proteína Desacopladora PDFbenito pereiraAún no hay calificaciones
- Nutricion 1Documento484 páginasNutricion 1PacoVivoAún no hay calificaciones
- Fosforilacion OxidativaDocumento17 páginasFosforilacion OxidativaChuy Huerta Lara100% (1)
- 3 Cadena Respiratoria y Fosforilacion OxidativaDocumento12 páginas3 Cadena Respiratoria y Fosforilacion OxidativaAngelina MerinoAún no hay calificaciones
- RESPIRACION. FinaldocxDocumento17 páginasRESPIRACION. FinaldocxlenfigueroaAún no hay calificaciones
- Resumen - Ciclo de KrebsDocumento5 páginasResumen - Ciclo de KrebsPamela Camarena TorresAún no hay calificaciones
- Práctica 4 Bombeo de Protones en LevaduraDocumento7 páginasPráctica 4 Bombeo de Protones en Levaduranatjeros12Aún no hay calificaciones
- Tema - 8 Los Orgánulos Celulares (II)Documento65 páginasTema - 8 Los Orgánulos Celulares (II)Clara NavarroAún no hay calificaciones
- Bioquímica Tema 2 (Fase Común Metabolismo)Documento27 páginasBioquímica Tema 2 (Fase Común Metabolismo)Karlos CaballeroAún no hay calificaciones
- Fosforilación Oxidativa Ciclo II-2023Documento22 páginasFosforilación Oxidativa Ciclo II-2023Ericson CastroAún no hay calificaciones
- 3 Discusión BioquímicaDocumento16 páginas3 Discusión BioquímicaMilver EscalanteAún no hay calificaciones
- Inhibidores Enzimaticos en La Cadena RespiratoriaDocumento5 páginasInhibidores Enzimaticos en La Cadena RespiratoriaFATIMA GERALDINE PIZAN OCUPAAún no hay calificaciones
- Biología Celular: Producto Parcial de Aprendizaje 3Documento164 páginasBiología Celular: Producto Parcial de Aprendizaje 3Oziel Moreno MoralesAún no hay calificaciones
- Fosforilación OxidativaDocumento7 páginasFosforilación Oxidativadarco16Aún no hay calificaciones