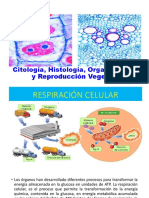
También podría gustarte
- 00010641397IA01S1109931010-CLASEBIOLOGIA2023-II (1)Documento51 páginas00010641397IA01S1109931010-CLASEBIOLOGIA2023-II (1)2325010006Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Metabolismo CelularDocumento35 páginasMetabolismo CelularJuan Carlos Ruiz Jimenez100% (1)
- Práctica N4 BioquimicaDocumento14 páginasPráctica N4 BioquimicaJose FernandoAún no hay calificaciones
- Obtención de Energía Por Los Organismos de La Madre TierraDocumento4 páginasObtención de Energía Por Los Organismos de La Madre TierraYola Marisol Condori Gutierrez94% (18)
- Informe de Respiracion CelularDocumento5 páginasInforme de Respiracion CelularJian marcos suriel corcinoAún no hay calificaciones
- Respiración CelularDocumento22 páginasRespiración CelularmarimarthaAún no hay calificaciones
- Catabolismo y AnabolismoDocumento27 páginasCatabolismo y AnabolismoJeimy Orantes100% (1)
- Composición Proteica de La Membrana PlasmáticaDocumento22 páginasComposición Proteica de La Membrana Plasmáticamonnn .-,2Aún no hay calificaciones
- Sesion 7 PPT - SincronoDocumento28 páginasSesion 7 PPT - SincronoInés Arias HuamaniAún no hay calificaciones
- MetabolismoDocumento27 páginasMetabolismoLibros Andrea100% (2)
- Glucolisis y Ciclo de KrebsDocumento11 páginasGlucolisis y Ciclo de KrebsanettedenissesgAún no hay calificaciones
- Cadena de Respiración CelularDocumento21 páginasCadena de Respiración CelularElizabeth BuestanAún no hay calificaciones
- Respiracion CelularDocumento8 páginasRespiracion CelularAndreaAún no hay calificaciones
- Respiración Celular: Proceso Vital para la VidaDocumento10 páginasRespiración Celular: Proceso Vital para la Vidasofia badilloAún no hay calificaciones
- Metabolismo Bioquimica PDFDocumento8 páginasMetabolismo Bioquimica PDFcarlos prudAún no hay calificaciones
- Resumito.2 Parcial. 2018.2Documento50 páginasResumito.2 Parcial. 2018.2Michelle CristineAún no hay calificaciones
- tema 9 y 10Documento9 páginastema 9 y 10carmencitahuecasAún no hay calificaciones
- BIOQUIMICADocumento17 páginasBIOQUIMICAIsabella1237Aún no hay calificaciones
- 2 Resumen Metabolismo Celular NEWDocumento11 páginas2 Resumen Metabolismo Celular NEWRonald Cajamarca RivadeneiraAún no hay calificaciones
- Colegio Adventista PimentelDocumento26 páginasColegio Adventista PimentelCamila RubioAún no hay calificaciones
- Tema 5. Metabolismo. Catabolismo y AnabolismoDocumento7 páginasTema 5. Metabolismo. Catabolismo y AnabolismoNatalia Milla LopezAún no hay calificaciones
- Grupo #5. Obtención de Energía Glucólisis y Respiración Celular NueeDocumento33 páginasGrupo #5. Obtención de Energía Glucólisis y Respiración Celular NueehgarciabonillaAún no hay calificaciones
- 1 MetabolismoDocumento34 páginas1 MetabolismoEspejel Torres Hakeem ZahidAún no hay calificaciones
- 3.4. Respiración CelularDocumento25 páginas3.4. Respiración CelularJaime PérezAún no hay calificaciones
- MetabolismoDocumento7 páginasMetabolismoFavian Fernandes100% (1)
- Transporte CelularDocumento9 páginasTransporte Celularsamueltareas122Aún no hay calificaciones
- Tema 8 Metabolismo Celular. BioenergéticaDocumento67 páginasTema 8 Metabolismo Celular. BioenergéticaGabriela DiazAún no hay calificaciones
- GLUCOLISISDocumento18 páginasGLUCOLISISTammi CardozhoAún no hay calificaciones
- Función de Nutrición - Relación y Reproducción CelularDocumento34 páginasFunción de Nutrición - Relación y Reproducción CelularDailys UtriaAún no hay calificaciones
- Expo Catabolismo y AnabolismoDocumento27 páginasExpo Catabolismo y AnabolismoMariana CCAún no hay calificaciones
- MetabolismoDocumento7 páginasMetabolismoFatima Serrano PerezAún no hay calificaciones
- TEMA 5 Respiracion Celular y Fermentacion.Documento5 páginasTEMA 5 Respiracion Celular y Fermentacion.mariaAún no hay calificaciones
- Área: Ciencias Naturales Grado: 7° Tema: La Respiración Es Una Actividad Catabólica, Respiración CelularDocumento31 páginasÁrea: Ciencias Naturales Grado: 7° Tema: La Respiración Es Una Actividad Catabólica, Respiración CelularYessica CordobaAún no hay calificaciones
- Metabolismo de carbohidratos: glucólisis, ciclo de Krebs y cadena respiratoriaDocumento15 páginasMetabolismo de carbohidratos: glucólisis, ciclo de Krebs y cadena respiratoriaCarolay ParedesAún no hay calificaciones
- Ciclo de Krebs - Rutas MetabólicasDocumento26 páginasCiclo de Krebs - Rutas MetabólicasJuan Daniel Dávalos CanteroAún no hay calificaciones
- Rutas MetabolicasDocumento49 páginasRutas Metabolicascralex22100% (1)
- Respiracion_celular_y_tp3Documento17 páginasRespiracion_celular_y_tp3tony3093Aún no hay calificaciones
- Equipo 10, Tema de Exposición.Documento24 páginasEquipo 10, Tema de Exposición.Fernanda IdkAún no hay calificaciones
- Semana 10. Obtención de Energía UPNDocumento35 páginasSemana 10. Obtención de Energía UPNcarmen marquinaAún no hay calificaciones
- Respiración CelularDocumento26 páginasRespiración CelularJoselin MuciñoAún no hay calificaciones
- Metabolismo Gluco Kbres MedicinaDocumento40 páginasMetabolismo Gluco Kbres MedicinaLuis eduardo La torre AdrianzénAún no hay calificaciones
- Tema8 MetabolismoDocumento18 páginasTema8 MetabolismoPablo SCAún no hay calificaciones
- Convergencia y Diversidad en El CatabolismoDocumento5 páginasConvergencia y Diversidad en El CatabolismojanansayaAún no hay calificaciones
- Biologia ExamenDocumento3 páginasBiologia Examendiego mendozaAún no hay calificaciones
- BiologiaDocumento8 páginasBiologiaDamian LovagniniAún no hay calificaciones
- Respiración CelularDocumento35 páginasRespiración CelularCharlin Cerna RuizAún no hay calificaciones
- Obtención de Energía A Través de La Degradación de CarbohidratosDocumento4 páginasObtención de Energía A Través de La Degradación de CarbohidratosIzel CachiAún no hay calificaciones
- Cuestionario 1. Distinga Entre Los Siguientes Conceptos: Oxidación de La Glucosa / Glucólisis / RespiraciónDocumento9 páginasCuestionario 1. Distinga Entre Los Siguientes Conceptos: Oxidación de La Glucosa / Glucólisis / RespiracióncarolinaAún no hay calificaciones
- Taller 3 BioquimicaDocumento11 páginasTaller 3 Bioquimicalinasofia6Aún no hay calificaciones
- Metabolismo CelularDocumento6 páginasMetabolismo CelularZoe DezalotAún no hay calificaciones
- Respiración Celular 2021Documento29 páginasRespiración Celular 202110HorasAún no hay calificaciones
- METABOLISMODocumento16 páginasMETABOLISMOCamila VillaAún no hay calificaciones
- Guía Respiracion CelularDocumento4 páginasGuía Respiracion CelularCristian Flores67% (3)
- Unidad 4. Metabolismo III PDFDocumento37 páginasUnidad 4. Metabolismo III PDFAdan MonterrosaAún no hay calificaciones
- Metabolismo (2)Documento41 páginasMetabolismo (2)ANGEL GABRIELAún no hay calificaciones
- Representación de Las Fases Relacionadas Con La RespiraciónDocumento3 páginasRepresentación de Las Fases Relacionadas Con La RespiraciónAfry ZuñigaAún no hay calificaciones
- METABOLISMODocumento16 páginasMETABOLISMODavid VelascoAún no hay calificaciones
- Clase 1 Farmacocinetica y FarmacodinamiaDocumento62 páginasClase 1 Farmacocinetica y FarmacodinamiaAlexa RomeroAún no hay calificaciones
- 04 Subsistema Medio Físico IDocumento11 páginas04 Subsistema Medio Físico ILuisc10Aún no hay calificaciones
- Anatomia (Sistema Tegumentario)Documento5 páginasAnatomia (Sistema Tegumentario)LESLY SIQUEY PEREZ SANCHEZAún no hay calificaciones
- Práctica #5 Balance HidricoDocumento12 páginasPráctica #5 Balance HidricoJOSE VALDEMAR OJEDA ROJASAún no hay calificaciones
- Resumen de Programas y ProyectosDocumento2 páginasResumen de Programas y ProyectosAmbiental InterventoriaAún no hay calificaciones
- Producción Microbiana de Ácido Hialurónico A Partir de Subproductos AgroindustrialesDocumento7 páginasProducción Microbiana de Ácido Hialurónico A Partir de Subproductos AgroindustrialesEsteban QuinteroAún no hay calificaciones
- Celda MicrobianaDocumento6 páginasCelda MicrobianaDavidAún no hay calificaciones
- TOMO I - Respiracion AikidoDocumento6 páginasTOMO I - Respiracion AikidoBrucewilsAún no hay calificaciones
- Informe de SaussureDocumento3 páginasInforme de SaussurePeach FancyAún no hay calificaciones
- 1 Protocolo de Ejercicios para Pacientes de Rehabilitación Cardiaca Con Patología IsquémicaDocumento12 páginas1 Protocolo de Ejercicios para Pacientes de Rehabilitación Cardiaca Con Patología IsquémicanilAún no hay calificaciones
- Procesos cognitivos básicos: Sensación y percepciónDocumento33 páginasProcesos cognitivos básicos: Sensación y percepciónROBERTO CARLOS VALDERRAMA MIRANDAAún no hay calificaciones
- Licopeno de Tomate 2 PDFDocumento6 páginasLicopeno de Tomate 2 PDFCarlo Jorge SepluvedaAún no hay calificaciones
- Aparato Digestivo (Propedeutica)Documento15 páginasAparato Digestivo (Propedeutica)america.fernanda.rocha.gomez.fmvzAún no hay calificaciones
- Uva VticDocumento62 páginasUva VticLuis Guardia EcheniqueAún no hay calificaciones
- EXAMEN GESTIÓN RESIDUOSDocumento2 páginasEXAMEN GESTIÓN RESIDUOSEddinson Jesus Panduro NoronhaAún no hay calificaciones
- Órganos articulatorios de la fonética españolaDocumento15 páginasÓrganos articulatorios de la fonética españolaDuda RochaAún no hay calificaciones
- Grupo3 Pino Tax - Impor.Documento14 páginasGrupo3 Pino Tax - Impor.CRISTIAN DE LA TORRE BENDEZUAún no hay calificaciones
- APF - Revista 2013Documento52 páginasAPF - Revista 2013Congreso Fitopatologia100% (1)
- Investigación Condicionamiento OperanteDocumento2 páginasInvestigación Condicionamiento Operanteandrea ochoaAún no hay calificaciones
- ENVIO 6 21 Dias para Activar El ADN Con Guillermo FerraraDocumento31 páginasENVIO 6 21 Dias para Activar El ADN Con Guillermo FerraraIsabelAcostaAún no hay calificaciones
- Arroz Dorado - Jorge MayerDocumento20 páginasArroz Dorado - Jorge MayerJorge MayerAún no hay calificaciones
- Hoja Seguridad Compuesto Rigido PVC Grado IndustrialDocumento7 páginasHoja Seguridad Compuesto Rigido PVC Grado IndustrialAlejandro HernandezAún no hay calificaciones
- Introduccion DermatoglifosDocumento2 páginasIntroduccion Dermatoglifosbolt169Aún no hay calificaciones
- Tarea 7 MorfopatoDocumento2 páginasTarea 7 MorfopatoJosé Manuel100% (2)
- Métodos InmunológicosDocumento19 páginasMétodos Inmunológicosmariipiii100% (3)
- PlacentaDocumento137 páginasPlacentaleodan riojas huamanAún no hay calificaciones
- Dte 15MDocumento8 páginasDte 15MGaby GonzalezAún no hay calificaciones
- 15-Guía Contracción Isometrica y DinamometrosDocumento4 páginas15-Guía Contracción Isometrica y DinamometrosCatalina WallisAún no hay calificaciones
- Neuroanatomia Clinica Snell 7a EdicionDocumento950 páginasNeuroanatomia Clinica Snell 7a EdicionMax Cr100% (2)
- Hermanito Hermanita Parte 1Documento54 páginasHermanito Hermanita Parte 1Yesenia Chambi LauraAún no hay calificaciones
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- La Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)De EverandLa Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)Calificación: 5 de 5 estrellas5/5 (4)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- DIETA MEDITERRANEA - Mejores Recetas de la Cocina Mediterranea Para Bajar de Peso SaludablementeDe EverandDIETA MEDITERRANEA - Mejores Recetas de la Cocina Mediterranea Para Bajar de Peso SaludablementeCalificación: 5 de 5 estrellas5/5 (4)
- Desastres y emergencias. Prevención, mitigación y preparaciónDe EverandDesastres y emergencias. Prevención, mitigación y preparaciónCalificación: 4 de 5 estrellas4/5 (5)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- La vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizDe EverandLa vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizCalificación: 5 de 5 estrellas5/5 (7)
- Teoría de juegos: Una introducción matemática a la toma de decisionesDe EverandTeoría de juegos: Una introducción matemática a la toma de decisionesCalificación: 4 de 5 estrellas4/5 (1)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Mega Piernas: Entrenamiento Científico de PiernasDe EverandMega Piernas: Entrenamiento Científico de PiernasCalificación: 4 de 5 estrellas4/5 (4)