También podría gustarte
- InjertosDocumento30 páginasInjertosFaby V. Rodríguez100% (1)
- Comunidades VegetalesDocumento88 páginasComunidades Vegetalesjcnmireles100% (2)
- Enfermedades en CitricosDocumento143 páginasEnfermedades en CitricosAdan Yomar FernándezAún no hay calificaciones
- Conceptos Basicos Agricultura 4623 CompletoDocumento20 páginasConceptos Basicos Agricultura 4623 CompletoLeandro GusmaoAún no hay calificaciones
- Las Lagrimas de NaraguyaDocumento13 páginasLas Lagrimas de NaraguyaRaulie GarciaAún no hay calificaciones
- FLOEMADocumento8 páginasFLOEMACarlos CarreteroAún no hay calificaciones
- MasoterapiaDocumento15 páginasMasoterapiaJerson Yovera Preciado100% (1)
- Laboratorio 2 ImbibicionDocumento18 páginasLaboratorio 2 ImbibicionEver Leonardo50% (2)
- Transporte de Agua en Las PlantasDocumento5 páginasTransporte de Agua en Las PlantasJuan Camilo C50% (2)
- Apertura y Cierre EstomáticoDocumento3 páginasApertura y Cierre EstomáticoAlieska Salazar CabreraAún no hay calificaciones
- 4.absorcion de Agua Por La RaizDocumento25 páginas4.absorcion de Agua Por La RaizHobie ArotomaAún no hay calificaciones
- Informe de Laboratorio TranspiracionDocumento13 páginasInforme de Laboratorio Transpiracionivan andres chagualo villareal50% (2)
- Edafologia N5Documento3 páginasEdafologia N5Abel Mauricio HuamánAún no hay calificaciones
- Existe Vegetación en El Terreno?Documento6 páginasExiste Vegetación en El Terreno?Steven SosaAún no hay calificaciones
- INF (1) Coloides EDAF 06Documento9 páginasINF (1) Coloides EDAF 06Edy Leon ChavezAún no hay calificaciones
- Inhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDocumento5 páginasInhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDarli Alvaro Camacho RojasAún no hay calificaciones
- El AporqueDocumento6 páginasEl AporqueMarcos LeonAún no hay calificaciones
- Tejido EpidermicoDocumento9 páginasTejido EpidermicoLuciana Sossiré Vásquez RegaladoAún no hay calificaciones
- Sistema Respiratorio - ConejoDocumento1 páginaSistema Respiratorio - ConejoAlberto GonzalezAún no hay calificaciones
- Valor Nutricional Del Esparrago (Asparagus Officinalis L.) para Uso Forrajero FINALDocumento28 páginasValor Nutricional Del Esparrago (Asparagus Officinalis L.) para Uso Forrajero FINALÁngel Miguel Cisneros SánchezAún no hay calificaciones
- Cuenca TrabajoDocumento36 páginasCuenca TrabajoGiuliana Sanchez MartinezAún no hay calificaciones
- Suelitos Materia Orgánica Marco TeoricoDocumento9 páginasSuelitos Materia Orgánica Marco TeoricoLuciaWarryAún no hay calificaciones
- Marco Teorico Tradiccion TranscripcionDocumento2 páginasMarco Teorico Tradiccion Transcripcionluis0% (2)
- Guía343 2020 - PRÁCTICA N°09Documento13 páginasGuía343 2020 - PRÁCTICA N°09Abel Mauricio Huamán100% (1)
- Edafologia WordDocumento15 páginasEdafologia WordCordova JasonAún no hay calificaciones
- Cierre Estomático y Ajuste OsmóticoDocumento5 páginasCierre Estomático y Ajuste OsmóticoCecilia Fernández JaramilloAún no hay calificaciones
- Encefalomielitis HemaglutinanteDocumento15 páginasEncefalomielitis HemaglutinanteJared JuarezAún no hay calificaciones
- Edafología - Aireación Del SueloDocumento46 páginasEdafología - Aireación Del SueloRoger Abel Huacal CortezAún no hay calificaciones
- 2 Informe Monolitos FinalDocumento17 páginas2 Informe Monolitos FinalWilliams Medina RamirezAún no hay calificaciones
- Macrofauna Del SueloDocumento6 páginasMacrofauna Del SuelovebtbrtbtrrnrnAún no hay calificaciones
- Reconocimiento de Células SanguíneasDocumento8 páginasReconocimiento de Células SanguíneasPaola Obando PadillaAún no hay calificaciones
- BIBLIOGRAFIA TexturaDocumento8 páginasBIBLIOGRAFIA TexturaCarla SemperiAún no hay calificaciones
- Factores Que Limitan El Crecimiento y Desarrollo de Las PlantasDocumento5 páginasFactores Que Limitan El Crecimiento y Desarrollo de Las PlantasPaul Gonzales VargasAún no hay calificaciones
- EdafizaciónDocumento6 páginasEdafizaciónRonald Valer PalaciosAún no hay calificaciones
- El Azufre en Plantas y SueloDocumento3 páginasEl Azufre en Plantas y SueloDisa PownedAún no hay calificaciones
- Forestación Práctica 5Documento4 páginasForestación Práctica 5infiltrado_1995Aún no hay calificaciones
- Germinacion de Las PlantasDocumento4 páginasGerminacion de Las PlantasNilton Steven Apaza BustinzaAún no hay calificaciones
- 15 PLANTAS-comprimidoDocumento15 páginas15 PLANTAS-comprimidocamila herreraAún no hay calificaciones
- El Humano Retorna Timidamente Al Bosque PDFDocumento157 páginasEl Humano Retorna Timidamente Al Bosque PDFAlexanderGilCampoAún no hay calificaciones
- Informe Salida de CampoDocumento24 páginasInforme Salida de Camposunda_hero0% (2)
- Plan de InvestigaciónDocumento3 páginasPlan de InvestigaciónRonald EscalanteAún no hay calificaciones
- Monografia Cultivo de ArvejaDocumento7 páginasMonografia Cultivo de ArvejaJhon Fredy Cuaran67% (3)
- CLASE 4. Componentes Externos Del TractorDocumento25 páginasCLASE 4. Componentes Externos Del TractorSeburanohAún no hay calificaciones
- Informe N°8 Nutrición MineralDocumento8 páginasInforme N°8 Nutrición MineralMishell B. Caballero ErazoAún no hay calificaciones
- Informe de Ciclo de ClvinDocumento2 páginasInforme de Ciclo de ClvinEstefani FloresAún no hay calificaciones
- 3.3. Factores Que Afectan Al Rendimiento FotosintéticoDocumento2 páginas3.3. Factores Que Afectan Al Rendimiento FotosintéticoJavier ZRAún no hay calificaciones
- Importancia de Las Plantas C4Documento6 páginasImportancia de Las Plantas C4EnriqueAún no hay calificaciones
- Ensayo Absorcion y TransporteDocumento2 páginasEnsayo Absorcion y TransporteAngie Assia100% (1)
- 9 AridisolesDocumento30 páginas9 AridisolesLaboratorio PumagoldAún no hay calificaciones
- Transpiracion - Semana 4Documento5 páginasTranspiracion - Semana 4Lizbeth Fiorella Rodriguez LujanAún no hay calificaciones
- 5 Bacterias FitopatógenasDocumento48 páginas5 Bacterias FitopatógenasJavier Valdivia100% (2)
- Informe de Fisiologia DifusionDocumento7 páginasInforme de Fisiologia DifusionJimmy Hurtado ValerioAún no hay calificaciones
- Latencia en SemillasDocumento6 páginasLatencia en SemillasBlanca Andrea CastilloAún no hay calificaciones
- Exposicion Agrotecnia Clase 1,2 3Documento109 páginasExposicion Agrotecnia Clase 1,2 3Mtsc Marco Desarrollo SostenibleAún no hay calificaciones
- Cuantificacion de Nematodos en El Suelo Por El Metodo de La BandejaDocumento17 páginasCuantificacion de Nematodos en El Suelo Por El Metodo de La BandejaFelicano Solano LimasAún no hay calificaciones
- ApioDocumento6 páginasApioLenUshioAún no hay calificaciones
- Crecimiento y DesarrolloDocumento6 páginasCrecimiento y DesarrolloAngel Ormeño Tananta0% (2)
- Experimento Proceso de Absorción de Color de Las FloresDocumento1 páginaExperimento Proceso de Absorción de Color de Las FloressjarguelleAún no hay calificaciones
- Ordenes de Los Suelos Según La USDADocumento4 páginasOrdenes de Los Suelos Según La USDACindy AndradeAún no hay calificaciones
- Informe Propagacion de Arboles FrutalesDocumento11 páginasInforme Propagacion de Arboles FrutalesAnamileth AguilarAún no hay calificaciones
- Fisiologia de Cultivos - Germinacion de Semilla 2Documento53 páginasFisiologia de Cultivos - Germinacion de Semilla 2Ray Kevin Leyva Garcia100% (1)
- Informe BocashiDocumento5 páginasInforme Bocashi123Aún no hay calificaciones
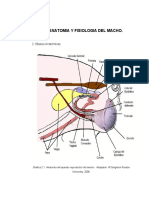
- Anatomia y Fisiologia Del MachoDocumento17 páginasAnatomia y Fisiologia Del MachoAleja Torres GaviriaAún no hay calificaciones
- Perfil de SueloDocumento3 páginasPerfil de SueloRomeo LópezAún no hay calificaciones
- La TranspiracionDocumento4 páginasLa Transpiracionxdelgado30100% (1)
- Anatomia de Los EstomasDocumento5 páginasAnatomia de Los EstomasNatt MejaiAún no hay calificaciones
- Rol de La Municipalidad en Agua y SaneamientoDocumento19 páginasRol de La Municipalidad en Agua y SaneamientoJerson Yovera Preciado100% (1)
- Formato de Informe Del Plan de Trabajo Guia 28Documento6 páginasFormato de Informe Del Plan de Trabajo Guia 28Jerson Yovera PreciadoAún no hay calificaciones
- Anexo #8 - MD-GUADALUPE - Formato de Informe de Plan de TrabajoDocumento3 páginasAnexo #8 - MD-GUADALUPE - Formato de Informe de Plan de TrabajoJerson Yovera PreciadoAún no hay calificaciones
- ZumoterapiaDocumento24 páginasZumoterapiaJerson Yovera PreciadoAún no hay calificaciones
- Función Fisiológica de Los Macronutrientes y Micronutrientes en Las PlantasDocumento4 páginasFunción Fisiológica de Los Macronutrientes y Micronutrientes en Las PlantasJerson Yovera Preciado33% (3)
- Evaluación Ambiental de La Bahía de PaitaDocumento4 páginasEvaluación Ambiental de La Bahía de PaitaJerson Yovera PreciadoAún no hay calificaciones
- Medio para Induccion de CallosDocumento5 páginasMedio para Induccion de CallosJerson Yovera PreciadoAún no hay calificaciones
- Conceptos de EspecieDocumento3 páginasConceptos de EspecieJerson Yovera PreciadoAún no hay calificaciones
- Conceptos de EspecieDocumento3 páginasConceptos de EspecieJerson Yovera PreciadoAún no hay calificaciones
- Chavi Mo ChicDocumento8 páginasChavi Mo ChicJerson Yovera PreciadoAún no hay calificaciones
- Haplotipos en Plantas CultivadasDocumento5 páginasHaplotipos en Plantas CultivadasJerson Yovera PreciadoAún no hay calificaciones
- Represión Catabolica Del Operón LacDocumento3 páginasRepresión Catabolica Del Operón LacJerson Yovera PreciadoAún no hay calificaciones
- Los ReflejosDocumento5 páginasLos ReflejosJerson Yovera PreciadoAún no hay calificaciones
- Propuesta de Manejo de Maguey Mezcalero en ViveroDocumento20 páginasPropuesta de Manejo de Maguey Mezcalero en Viveroprimitivo100% (1)
- Atropellamiento de Fauna en Rutas y Caminos Internos Del Parque Nacional Iguazú, MisionesDocumento13 páginasAtropellamiento de Fauna en Rutas y Caminos Internos Del Parque Nacional Iguazú, MisionesBiblioteca Perito Francisco P. Moreno.Argentina100% (2)
- Prácticas Vegetativas y Agronómicas PDFDocumento25 páginasPrácticas Vegetativas y Agronómicas PDFPatty RossalAún no hay calificaciones
- Evaluación Del Habito de Alimentación de La Larva de Plutella Xylostella (L.) (Lepidoptera: Plutellidae)Documento44 páginasEvaluación Del Habito de Alimentación de La Larva de Plutella Xylostella (L.) (Lepidoptera: Plutellidae)Pao Becerra E0% (1)
- Explique Que Son Los CarbohidratosDocumento2 páginasExplique Que Son Los CarbohidratosJosh Tapia MuciñoAún no hay calificaciones
- Me Divierto Con Las FloresDocumento5 páginasMe Divierto Con Las FloresJaime Abel Castillo AvilaAún no hay calificaciones
- Historia de La Exploracion Botanica de La Provincia de Albacete-1Documento21 páginasHistoria de La Exploracion Botanica de La Provincia de Albacete-1drivera789Aún no hay calificaciones
- Madera Tema 9Documento61 páginasMadera Tema 9Cristhian PLAún no hay calificaciones
- Ii Corto Biologia 9Documento2 páginasIi Corto Biologia 9LIZETH0% (1)
- Arboles Utilizados para Madera PreciosaDocumento9 páginasArboles Utilizados para Madera PreciosaMelvin CruzAún no hay calificaciones
- HerbarioDocumento6 páginasHerbarioRRAún no hay calificaciones
- PRACTICA Nº6 CELULA VegetalDocumento5 páginasPRACTICA Nº6 CELULA VegetalAnonymous HznQwKPk1Aún no hay calificaciones
- Efectos de La Salinidad y La Sodicidad en Los CultivosDocumento7 páginasEfectos de La Salinidad y La Sodicidad en Los CultivosJorge Vilela DuranAún no hay calificaciones
- CALENDARIO COMUNAL Capitan Evarisro AmesquitaDocumento9 páginasCALENDARIO COMUNAL Capitan Evarisro Amesquitanestor_aquinoAún no hay calificaciones
- Plantas AcuaticasDocumento14 páginasPlantas AcuaticasFernandoChiAún no hay calificaciones
- Técnicas Le Almacenaiiento Y Conseruición Le Frutas' Horjaulas FrescasDocumento142 páginasTécnicas Le Almacenaiiento Y Conseruición Le Frutas' Horjaulas FrescasRina Valera VasquezAún no hay calificaciones
- Av. Teoria y PracticaDocumento111 páginasAv. Teoria y PracticaDJUga2Aún no hay calificaciones
- La SabanaDocumento7 páginasLa SabanaLady SanchezAún no hay calificaciones
- ESTUDIO DE BIODIVERSIDAD EN EL CACAO - CONSERVACION & DESARROLLO, Make A Donation@ccd - Org.ec / Haga Una DonaciónDocumento92 páginasESTUDIO DE BIODIVERSIDAD EN EL CACAO - CONSERVACION & DESARROLLO, Make A Donation@ccd - Org.ec / Haga Una DonaciónCONSERVACION Y DESARROLLO SOSTENIBLEAún no hay calificaciones
- Visista Tecnica A Un AserraderoDocumento11 páginasVisista Tecnica A Un AserraderoArnold Alexander Campos Rivera100% (1)
- Informe Pigmentos FotosinteticosDocumento5 páginasInforme Pigmentos FotosinteticosGonzalo PortillaAún no hay calificaciones
- INFORME DE TESIS EN AYMARA 4to PDFDocumento93 páginasINFORME DE TESIS EN AYMARA 4to PDFRaul Antonio Ramirez100% (1)
- 1005 Dinamicas GrupoDocumento22 páginas1005 Dinamicas GrupoPuccayouAún no hay calificaciones
- VoldosDocumento367 páginasVoldosDaniel ComeglioAún no hay calificaciones
- Cuerpo Del ProyectoDocumento52 páginasCuerpo Del ProyectoDāvīd TêdAún no hay calificaciones