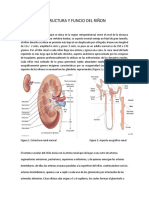
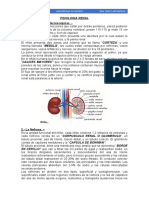
También podría gustarte
- Instituto Superior Tecnológico Cruz Roja EcuatorianaDocumento7 páginasInstituto Superior Tecnológico Cruz Roja EcuatorianaAngelicaEmoxaAún no hay calificaciones
- Material de Apoyo Al Estudio Del Aparato UrinarioDocumento19 páginasMaterial de Apoyo Al Estudio Del Aparato UrinarioEsmeraldaAún no hay calificaciones
- Aparato Urinario Marco TeòricoDocumento18 páginasAparato Urinario Marco TeòricoAlxAgorAún no hay calificaciones
- Practica No12 Aparato Urinario.Documento15 páginasPractica No12 Aparato Urinario.Karina Ambriz ParedesAún no hay calificaciones
- Aparato UrinarioDocumento11 páginasAparato UrinarioDariiana Benavides100% (1)
- Resumen UrinarioDocumento5 páginasResumen UrinarioValeria Belén SantanderAún no hay calificaciones
- Apuntes Nefro (Principal)Documento120 páginasApuntes Nefro (Principal)Crixalis 145Aún no hay calificaciones
- Estructura de Los RiñonesDocumento16 páginasEstructura de Los RiñonesKathy AmoradiosAún no hay calificaciones
- Anatomía Del RiñónDocumento4 páginasAnatomía Del RiñónMaite PashmayAún no hay calificaciones
- Sistema UrinarioDocumento8 páginasSistema UrinarioAlexis S. RuizAún no hay calificaciones
- Nefrectomia, Trabajo FinalDocumento15 páginasNefrectomia, Trabajo FinalJose Lopez De [OssioAún no hay calificaciones
- Anatomia y Fisiologia Renal para EnfermerasDocumento172 páginasAnatomia y Fisiologia Renal para EnfermerasMaria Camila Ramírez Gutiérrez100% (1)
- Tema 4 AP. Urinario AnatomíaDocumento11 páginasTema 4 AP. Urinario AnatomíaJuanjo MedinaAún no hay calificaciones
- RIÑÓNDocumento7 páginasRIÑÓNParker GreenAún no hay calificaciones
- RiñónDocumento14 páginasRiñónJose Alberto Lazcano BrunoAún no hay calificaciones
- Sistema Urinario Histologia Uabp 1Documento7 páginasSistema Urinario Histologia Uabp 1lua robertiAún no hay calificaciones
- Histología Del RiñonDocumento6 páginasHistología Del RiñonKarime LopezavilaAún no hay calificaciones
- NefrologíaDocumento59 páginasNefrologíakarlasifontesAún no hay calificaciones
- Tejido MuscularDocumento9 páginasTejido MuscularDaniela HeLoAún no hay calificaciones
- Cs. Biológicas - Sist. RenalDocumento59 páginasCs. Biológicas - Sist. RenalAna belen GomezAún no hay calificaciones
- NEFRONADocumento14 páginasNEFRONANadia Sabine Soler AlarcónAún no hay calificaciones
- Copia de Portada Tesis Tercer SemestreDocumento19 páginasCopia de Portada Tesis Tercer SemestreMariana RoblesAún no hay calificaciones
- Fisiología RenalDocumento20 páginasFisiología RenalAngel GUAún no hay calificaciones
- RIÑONESDocumento9 páginasRIÑONESEstibaliz Carmona BerriosAún no hay calificaciones
- Morfofisiologia Humana IIDocumento18 páginasMorfofisiologia Humana IIDanna TorrellesAún no hay calificaciones
- Aparato UrinarioDocumento64 páginasAparato UrinarioSalvador BARRON MELOAún no hay calificaciones
- Histologia - Resumen de Aparato UrinarioDocumento11 páginasHistologia - Resumen de Aparato UrinarioValeria MéndezAún no hay calificaciones
- Fisiologia y Fisiopatologia RenalDocumento20 páginasFisiologia y Fisiopatologia RenalAnnibel Flores ReyesAún no hay calificaciones
- Estructura y Funcio Del RiñonDocumento4 páginasEstructura y Funcio Del RiñonKEILA INTERNETAún no hay calificaciones
- La Nefrona, Tubulos Proximales y DistalesDocumento2 páginasLa Nefrona, Tubulos Proximales y DistalesHeidi Esther BaezAún no hay calificaciones
- Tarea 2 Primer Parcial UroanalisisDocumento8 páginasTarea 2 Primer Parcial UroanalisisAmdres AlmonteAún no hay calificaciones
- Apuntes Aparato UrinarioDocumento12 páginasApuntes Aparato UrinarioPaz GarciaAún no hay calificaciones
- Fisiologia RenalDocumento20 páginasFisiologia RenalMonique GloverAún no hay calificaciones
- Anatomia Y Fisiología Del Sistema Renal: Características GeneralesDocumento15 páginasAnatomia Y Fisiología Del Sistema Renal: Características GeneralesgomezAún no hay calificaciones
- Fisiologia RenalDocumento21 páginasFisiologia RenalDayana Ruiz HerreraAún no hay calificaciones
- Aparato Urinario 199910 DownloableDocumento8 páginasAparato Urinario 199910 DownloableFrank FQAún no hay calificaciones
- Aparato Urinario 199910 DownloableDocumento8 páginasAparato Urinario 199910 DownloableFrank FQAún no hay calificaciones
- Capitulo Ii RiñonDocumento36 páginasCapitulo Ii Riñonadrianamargarita0007.amooAún no hay calificaciones
- AnatomíaDocumento66 páginasAnatomíaMarìa Daniela ZambranoAún no hay calificaciones
- Tema 10. Aparato UrinarioDocumento14 páginasTema 10. Aparato UrinarioanahitamarAún no hay calificaciones
- Los RiñonesDocumento8 páginasLos RiñonesPedro Jose Felipa BazalarAún no hay calificaciones
- Anatomía y Fisiología Del Riñón, Formación de Orina, Equilibrio Hidroelectrolític0 y Ácido BaseDocumento19 páginasAnatomía y Fisiología Del Riñón, Formación de Orina, Equilibrio Hidroelectrolític0 y Ácido BasejllontopllaAún no hay calificaciones
- Anatomia de Los RiñonesDocumento6 páginasAnatomia de Los RiñonesAlexandre QuisbertAún no hay calificaciones
- Elementos de La Función RenalDocumento8 páginasElementos de La Función RenalFlorencia BarsiAún no hay calificaciones
- Generalidades de Los RiñonesDocumento12 páginasGeneralidades de Los RiñonesMariaAlexandraPinoAún no hay calificaciones
- CURSODEGENITOURINARIO ManualDocumento80 páginasCURSODEGENITOURINARIO Manualcampsar50% (2)
- Principal UroDocumento253 páginasPrincipal UroAriel GuerreroAún no hay calificaciones
- ANATOMIADocumento6 páginasANATOMIAMontserrat ZaldoAún no hay calificaciones
- Unidad 6. El RiñonDocumento10 páginasUnidad 6. El RiñonFederico Pennacchioni polaAún no hay calificaciones
- Histologia Riñones PuntosDocumento13 páginasHistologia Riñones Puntosgabriela escobarAún no hay calificaciones
- Clase de RenalDocumento34 páginasClase de RenalDeniseDelPuertoAún no hay calificaciones
- Apuntes BiologiaDocumento4 páginasApuntes BiologiaMartin Paredes SanchezAún no hay calificaciones
- Anatomia y Fisiologia Renal Del Riñon - ResumenDocumento8 páginasAnatomia y Fisiologia Renal Del Riñon - ResumenEstephany MolinaAún no hay calificaciones
- Patologías Del Aparato Genito-Urinario PDFDocumento94 páginasPatologías Del Aparato Genito-Urinario PDFEster SadornilAún no hay calificaciones
- Fisiologia RenalDocumento13 páginasFisiologia RenalDelmira Andia challcoAún no hay calificaciones
- Tema 10. Anatomía y Fisología Del Aparato ExcretorDocumento15 páginasTema 10. Anatomía y Fisología Del Aparato Excretorlulu-luchiAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Programación 2010A Fisiología Grupo 03Documento7 páginasProgramación 2010A Fisiología Grupo 03ehtoledoc2127Aún no hay calificaciones
- Programación 2010A Fisiología Grupo 01Documento7 páginasProgramación 2010A Fisiología Grupo 01ehtoledoc2127Aún no hay calificaciones
- Fisiología Del Aparato RespiratorioDocumento15 páginasFisiología Del Aparato Respiratorioehtoledoc212767% (3)
- Programación 2010A Fisiología Grupo 01Documento7 páginasProgramación 2010A Fisiología Grupo 01ehtoledoc2127Aún no hay calificaciones
- Programación 2010A Farmacología Grupo 01Documento5 páginasProgramación 2010A Farmacología Grupo 01ehtoledoc21270% (1)
- Sistema Circulatorio Corazón y MiaDocumento12 páginasSistema Circulatorio Corazón y Miaehtoledoc100% (8)
- Sistema Circulatorio Corazón y MiaDocumento12 páginasSistema Circulatorio Corazón y Miaehtoledoc100% (8)
- 2009 Grupos y Acciones FarmacológicosDocumento12 páginas2009 Grupos y Acciones Farmacológicosehtoledoc212783% (6)
- Fisiología de Contracción MuscularDocumento7 páginasFisiología de Contracción Muscularehtoledoc212792% (12)
- Fisiología ÓseaDocumento4 páginasFisiología Óseaehtoledoc212775% (4)
- 4 Vías de Administración FarmacológicaDocumento55 páginas4 Vías de Administración Farmacológicaehtoledoc212792% (12)
- 2 Formas FarmaceúticasDocumento39 páginas2 Formas Farmaceúticasehtoledoc2127100% (5)
- Represent Antes Del MercantilismoDocumento12 páginasRepresent Antes Del MercantilismoAnDy FiTzAún no hay calificaciones
- La Zona de Reclamacion Guayana EsequibaDocumento10 páginasLa Zona de Reclamacion Guayana EsequibamabeAún no hay calificaciones
- Arteterapia y EmocionesDocumento9 páginasArteterapia y EmocionesFauna Tiva100% (8)
- Cucuta Verde y Exuberante Nov.2013Documento66 páginasCucuta Verde y Exuberante Nov.2013Mauricio Rozo100% (2)
- Sesión CeroDocumento11 páginasSesión CeroFatyma OzunaAún no hay calificaciones
- Diagrama de Flujo para La Elaboracion Camiseta PoloDocumento2 páginasDiagrama de Flujo para La Elaboracion Camiseta Pologalaxy elastronautaAún no hay calificaciones
- La Carta NotarialDocumento2 páginasLa Carta NotarialEdgar Cayllahua QuispeAún no hay calificaciones
- Plan Conceptual - ModificadoDocumento13 páginasPlan Conceptual - ModificadoHerman Diaz RomeroAún no hay calificaciones
- Propulsion Electrica de BuquesDocumento17 páginasPropulsion Electrica de Buqueslyndonpintado100% (2)
- LABORATORIO3Documento20 páginasLABORATORIO3edison guarangoAún no hay calificaciones
- Guerra Del PacificoDocumento60 páginasGuerra Del PacificoAngel Hassan Suarez CuellarAún no hay calificaciones
- Personalidad Adictiva-PresentacionDocumento7 páginasPersonalidad Adictiva-Presentacionsergio gonzalezAún no hay calificaciones
- Prevención de Factores de Riesgo Psicosociales en Enfermeras. InvestigaciónDocumento80 páginasPrevención de Factores de Riesgo Psicosociales en Enfermeras. InvestigaciónJavier ZúñigaAún no hay calificaciones
- Politica 5sDocumento2 páginasPolitica 5sADRIANYI67% (3)
- Proyecto Formativo Ventas de Productos y Ss Grado 10 A.H.G. 352133Documento8 páginasProyecto Formativo Ventas de Productos y Ss Grado 10 A.H.G. 352133Michael Steven Diaz Morales100% (3)
- 126-156 Número DecimalDocumento7 páginas126-156 Número DecimalBasted MDAún no hay calificaciones
- Normativa D S N 227 Reglamento Clasificacion Hotel Motel y AparthotelDocumento13 páginasNormativa D S N 227 Reglamento Clasificacion Hotel Motel y AparthotelAlexa AleAún no hay calificaciones
- La Mala Convivencia en El AulaDocumento2 páginasLa Mala Convivencia en El Aulajames gonzales pachecoAún no hay calificaciones
- Dinámica de Grupos-Definición de GruposDocumento30 páginasDinámica de Grupos-Definición de GruposgeovannirsAún no hay calificaciones
- Trabajo de Maquinas Termicas Refrigerador DomesticoDocumento11 páginasTrabajo de Maquinas Termicas Refrigerador Domesticojhony galindo aumaAún no hay calificaciones
- Universidad Nacional de San Agustin SupervicionDocumento5 páginasUniversidad Nacional de San Agustin SupervicionDan PillcoAún no hay calificaciones
- Diapositivas de Ambito EspacialDocumento9 páginasDiapositivas de Ambito EspacialAle Queen100% (1)
- Ensayo Geopolítico Del Potencial de La Energía Solar Fotovoltaica Que Contribuya Al Desarrollo Sostenible de BoliviaDocumento10 páginasEnsayo Geopolítico Del Potencial de La Energía Solar Fotovoltaica Que Contribuya Al Desarrollo Sostenible de BoliviaFranz Carlos Conalde TejerinaAún no hay calificaciones
- Construcción y Montaje de Buzones de Carguío en Minería (Parte1)Documento41 páginasConstrucción y Montaje de Buzones de Carguío en Minería (Parte1)David Caballero Leaño100% (1)
- Biografia Daniel GolemanDocumento3 páginasBiografia Daniel GolemanRafa Montenegro Saavedra100% (2)
- Informe N 034 Vigilancia A LicoreeriasDocumento1 páginaInforme N 034 Vigilancia A LicoreeriasLUISAún no hay calificaciones
- Los Vicarios Parroquiales en El Código de Derecho CanónicoDocumento2 páginasLos Vicarios Parroquiales en El Código de Derecho CanónicoFidel SanchezAún no hay calificaciones
- Libro Mov. Cta. CorrienteDocumento1 páginaLibro Mov. Cta. CorrienteMiguel H JAún no hay calificaciones
- Auditoria FinancieraDocumento8 páginasAuditoria FinancieraAbel RamírezAún no hay calificaciones
- Hada MadrinaDocumento13 páginasHada MadrinaVanesa Anderson FyhnAún no hay calificaciones