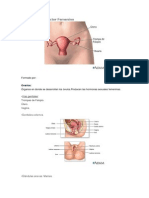
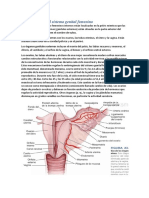
También podría gustarte
- Anatomía y fisiología de la mamaDocumento33 páginasAnatomía y fisiología de la mamaMauRodriguezCardoso100% (2)
- Glándulas MamariasDocumento5 páginasGlándulas MamariasRandy Reina RiveroAún no hay calificaciones
- Glandula MamariaDocumento6 páginasGlandula MamariaGisela MartinezAún no hay calificaciones
- Anatomia y Fisiologia de La Glandula Ma Maria WordDocumento6 páginasAnatomia y Fisiologia de La Glandula Ma Maria WordWilderAún no hay calificaciones
- Anatomía y fisiología de la glándula mamariaDocumento13 páginasAnatomía y fisiología de la glándula mamariaKriss David DelgadoAún no hay calificaciones
- EMBRIOLOG+ìA, HISTOLOG+ìA Y ANATOM+ìA DE LA MAMADocumento9 páginasEMBRIOLOG+ìA, HISTOLOG+ìA Y ANATOM+ìA DE LA MAMAoscarydalcyAún no hay calificaciones
- Histologia Normal de La Glándula MamariaDocumento10 páginasHistologia Normal de La Glándula MamariaLid YungAún no hay calificaciones
- Glandula Mamaria - HistologiaDocumento2 páginasGlandula Mamaria - HistologiaLuz JerezAún no hay calificaciones
- MamasDocumento19 páginasMamasvidalma RodriguezAún no hay calificaciones
- Anatomia y Fisiologia de La Glandula MamariaDocumento13 páginasAnatomia y Fisiologia de La Glandula MamariaMARYVONAún no hay calificaciones
- Anatomia y Fisiologia de La Glandula MamDocumento9 páginasAnatomia y Fisiologia de La Glandula MamLizeth MenaAún no hay calificaciones
- Apa. Reproductor FemeninoDocumento83 páginasApa. Reproductor FemeninoGeeder NohAún no hay calificaciones
- Embriologia de la mama y anatomía funcionalDocumento19 páginasEmbriologia de la mama y anatomía funcionalJose Luis SaldañaAún no hay calificaciones
- Histologia Ovarios y TrompasDocumento13 páginasHistologia Ovarios y TrompasMorenaAún no hay calificaciones
- HistologíaDocumento3 páginasHistologíaNancyGonzalezAún no hay calificaciones
- Glandulas MamariasDocumento4 páginasGlandulas MamariasYelitza GarceteAún no hay calificaciones
- Apuntes Patol Benigna 2013-Zp-rzDocumento53 páginasApuntes Patol Benigna 2013-Zp-rzCarlos Castro InzunzaAún no hay calificaciones
- GLANDULA-MAMARIA-obstetriciaDocumento21 páginasGLANDULA-MAMARIA-obstetriciaLuis Anderson Lozada GonzalezAún no hay calificaciones
- Anatomía e histología de la mamaDocumento3 páginasAnatomía e histología de la mamaDavid Clemo SteelAún no hay calificaciones
- Glandula Mamaria MorfofuncionDocumento8 páginasGlandula Mamaria MorfofuncionAnthony ZuritaAún no hay calificaciones
- Anatómica de La MamaDocumento9 páginasAnatómica de La MamaSusanValderaSimbronAún no hay calificaciones
- Anatomia y Fisiologia de La Glandula MamariaDocumento7 páginasAnatomia y Fisiologia de La Glandula MamariaCarmen Rosa Ortuño PachecoAún no hay calificaciones
- Anatomía de la mama femeninaDocumento4 páginasAnatomía de la mama femeninaFRANCISCO MONTAÑOAún no hay calificaciones
- Anatomía Histologa Glandula MamariaDocumento6 páginasAnatomía Histologa Glandula MamariaRogerio TavaresAún no hay calificaciones
- Resumen de Glandulas MamariasDocumento5 páginasResumen de Glandulas MamariasKatherine.acunaariasAún no hay calificaciones
- Glandulas MamariasDocumento8 páginasGlandulas MamariasEstefany SuárezAún no hay calificaciones
- Glándula Mamaria 1Documento3 páginasGlándula Mamaria 1XiomaraDayannaBancesRamirezAún no hay calificaciones
- Glandulas MamariasDocumento56 páginasGlandulas MamariasEfraín TelloAún no hay calificaciones
- Anatomía y FisiologiaDocumento5 páginasAnatomía y FisiologiaALISON MAYA ZARATE RODRIGUEZAún no hay calificaciones
- Anatomia de La MamaDocumento15 páginasAnatomia de La MamaWilder83% (6)
- Aparato Reproductor FemeninoDocumento62 páginasAparato Reproductor FemeninoMvm LakmirAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento23 páginasAparato Reproductor FemeninoMaritza Lobos HernandezAún no hay calificaciones
- Aparato Reproductor Femenino PresentacionDocumento44 páginasAparato Reproductor Femenino PresentacionEmily Velasquez100% (1)
- Clase 2 Anatomía e Histología Normal de La Mama 2020Documento55 páginasClase 2 Anatomía e Histología Normal de La Mama 2020FranciscaqcAún no hay calificaciones
- Anatomía Materna AplicadaDocumento6 páginasAnatomía Materna AplicadaAlessandro MoralesAún no hay calificaciones
- Anatomía de la glándula mamariaDocumento2 páginasAnatomía de la glándula mamariaAna MalaisiAún no hay calificaciones
- SIS Reproductor TallerDocumento15 páginasSIS Reproductor TallerVivian ParragaAún no hay calificaciones
- Anatomía testículoDocumento8 páginasAnatomía testículoBrenda Lizet Hernandez SandovalAún no hay calificaciones
- Generalidades Del Sistema Genital FemeninoDocumento13 páginasGeneralidades Del Sistema Genital FemeninoFede AranaAún no hay calificaciones
- Histologia Del Sistema Reproductor FemenidoDocumento17 páginasHistologia Del Sistema Reproductor FemenidoRoslenny GrullonAún no hay calificaciones
- Manual Lactancia Materna-62-74Documento13 páginasManual Lactancia Materna-62-74Esteban Angel SantanderAún no hay calificaciones
- Generalidades Del Aparato Genital Femenino OvarioDocumento53 páginasGeneralidades Del Aparato Genital Femenino OvarioLuis Martinez100% (1)
- Arf TeoriaDocumento1 páginaArf TeoriaRiver MoralesAún no hay calificaciones
- Sistema ReproductorDocumento17 páginasSistema ReproductorGianna ManziAún no hay calificaciones
- Tema A: Anatomía Materna e Infantil en Lactancia MaternaDocumento20 páginasTema A: Anatomía Materna e Infantil en Lactancia MaternaInma PablosAún no hay calificaciones
- Histologia de Mama.Documento2 páginasHistologia de Mama.Aylin Antonella Jacobi RohrAún no hay calificaciones
- Sistema reproductor femeninoDocumento46 páginasSistema reproductor femeninoSANTIAGO AGUIRRE GIRALDOAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento13 páginasAparato Reproductor FemeninoNair MartínezAún no hay calificaciones
- GINECOLOGÍA NasanielDocumento30 páginasGINECOLOGÍA NasanielNasaniele RodríguezAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento38 páginasAparato Reproductor FemeninoYaribel Kelly100% (1)
- Mehu604 - U1 - S15 - MAMA - TEORIADocumento23 páginasMehu604 - U1 - S15 - MAMA - TEORIAGESU PIERO LOPEZ MEJIAAún no hay calificaciones
- Anatomia de La Mama - CorrelacionDocumento3 páginasAnatomia de La Mama - CorrelacionAndrea Ruiz SolanoAún no hay calificaciones
- El Sistema Reproductor Masculino y FemeninoDocumento22 páginasEl Sistema Reproductor Masculino y FemeninoMeninet San BenitoAún no hay calificaciones
- Anatomía de Las MamasDocumento8 páginasAnatomía de Las MamasArianna carvajalAún no hay calificaciones
- Cancer de MamaaDocumento24 páginasCancer de MamaaCarlos BossaAún no hay calificaciones
- Histología MamaDocumento9 páginasHistología MamaDenia Ortega100% (1)
- Aparato Reproductor Femenino Daniel CaamañoDocumento15 páginasAparato Reproductor Femenino Daniel CaamañoDaniel CRAún no hay calificaciones
- 29-Aparato Reproductor FemeninoDocumento13 páginas29-Aparato Reproductor FemeninoShayAlarcónAún no hay calificaciones
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Up3 NutricionDocumento7 páginasUp3 Nutriciontiziana.batistuttiAún no hay calificaciones
- Resumen Nutri Up3Documento4 páginasResumen Nutri Up3tiziana.batistuttiAún no hay calificaciones
- Guía Didáctica UP3Documento4 páginasGuía Didáctica UP3tiziana.batistuttiAún no hay calificaciones
- 2 - UP 1.2 Respaso Estructura Glucidos y LipidosDocumento28 páginas2 - UP 1.2 Respaso Estructura Glucidos y Lipidostiziana.batistuttiAún no hay calificaciones
- Guía Didáctica UP2 SexualidadDocumento5 páginasGuía Didáctica UP2 SexualidadMicaelle AlbuquerqueAún no hay calificaciones
- Informe Up1 NutricionDocumento1 páginaInforme Up1 Nutriciontiziana.batistuttiAún no hay calificaciones
- Up2 NutriDocumento10 páginasUp2 Nutritiziana.batistuttiAún no hay calificaciones
- Fisio Up2Documento11 páginasFisio Up2tiziana.batistuttiAún no hay calificaciones
- Mellizos y Gemelos Editable WorksDocumento8 páginasMellizos y Gemelos Editable WorksThalia FloresAún no hay calificaciones
- Sistema Endocrino IIDocumento15 páginasSistema Endocrino IIpaz BnstrAún no hay calificaciones
- Esteroides Anabolizantes AndrogénicosDocumento50 páginasEsteroides Anabolizantes AndrogénicosMarc Mier Cervantes100% (1)
- VULNERABILIDADDocumento35 páginasVULNERABILIDADlaura acostaAún no hay calificaciones
- Climaterio y MenopausiaDocumento7 páginasClimaterio y MenopausiaJose Enrique Cruz Moreno100% (1)
- CRECIMIENOT Y DESARROLLO DEL ADOLESCENTEDocumento30 páginasCRECIMIENOT Y DESARROLLO DEL ADOLESCENTEMaydelin GuzmánAún no hay calificaciones
- 1.audesirk - 2013 - Endoc-745-751Documento7 páginas1.audesirk - 2013 - Endoc-745-751J MAún no hay calificaciones
- Los Disruptores EndocrinosDocumento6 páginasLos Disruptores EndocrinosJose Antonio Quintero Recio100% (1)
- Mapa Conceptual Piel Tarea 1Documento2 páginasMapa Conceptual Piel Tarea 1NandaAún no hay calificaciones
- Uso de Anabólicos en BovinosDocumento12 páginasUso de Anabólicos en BovinosCarlos Ruben BlancoAún no hay calificaciones
- Unidad 11. GonadasDocumento279 páginasUnidad 11. GonadasMichelle OlánAún no hay calificaciones
- Cuestionario Práctica 1 QS Glucosa Sérica y CDocumento3 páginasCuestionario Práctica 1 QS Glucosa Sérica y CDiana HernandezAún no hay calificaciones
- Mecanica de FluidosDocumento9 páginasMecanica de FluidosAnggie Nathalie Tobar ContrerasAún no hay calificaciones
- Ciclo EndometrialDocumento10 páginasCiclo EndometrialAGUILAR LUISAún no hay calificaciones
- 3 P Octavo Preguntas Icfes SemanalesDocumento13 páginas3 P Octavo Preguntas Icfes SemanalesPaola Ruiz CardenasAún no hay calificaciones
- Tema 10 Sistema Urogenital IDocumento34 páginasTema 10 Sistema Urogenital IKeydellAún no hay calificaciones
- Banco de Preguntas de Sistema RespiratorioDocumento6 páginasBanco de Preguntas de Sistema RespiratorioFelipe Ignacio Gutierrez BarriosAún no hay calificaciones
- Lesiones de Hipófisis y AbordajesDocumento45 páginasLesiones de Hipófisis y AbordajesGerardo Osorio Leon100% (1)
- Corticoesteroides: glucocorticoides y mineralocorticoidesDocumento82 páginasCorticoesteroides: glucocorticoides y mineralocorticoidesWilfredo CastilloAún no hay calificaciones
- Borrador de Informe Final de Tesina - Estres LaboralDocumento95 páginasBorrador de Informe Final de Tesina - Estres LaboralKarina TevezAún no hay calificaciones
- Cetoacidosis Diabética en PediatriaDocumento21 páginasCetoacidosis Diabética en PediatriaJeiner HernandezAún no hay calificaciones
- Trastornos MenstrualesDocumento16 páginasTrastornos MenstrualesDaniela MéndezAún no hay calificaciones
- Función Endocrina AlteracionesDocumento26 páginasFunción Endocrina AlteracionesJesús SalazarAún no hay calificaciones
- SPMDocumento18 páginasSPMMilagros PichardoAún no hay calificaciones
- Aparato reproductorDocumento5 páginasAparato reproductorSamuel PozAún no hay calificaciones
- AmenorreaDocumento16 páginasAmenorreaFelipe GiraldoAún no hay calificaciones
- Revisar Glosario, Bibliografia, Cap. Anex-1Documento7 páginasRevisar Glosario, Bibliografia, Cap. Anex-1Romero EspinozaAún no hay calificaciones
- Alba-Ppt-Órganos-SexualesDocumento42 páginasAlba-Ppt-Órganos-SexualesSenddey Maciel MagañaAún no hay calificaciones
- EmbriologiaDocumento5 páginasEmbriologiaJhey Kenider Benites IzquierdoAún no hay calificaciones
- Diabetes Emotiva: Mito o VerdadDocumento2 páginasDiabetes Emotiva: Mito o VerdadMartha Paola Arellano Salazar100% (2)