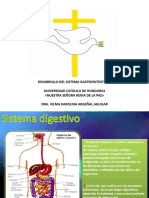
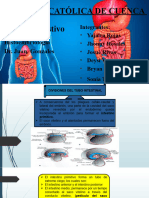
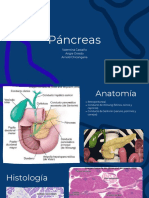
También podría gustarte
- Aparato DigestivoDocumento10 páginasAparato DigestivoKamila Castillo VegaAún no hay calificaciones
- Aparato Digestivo Embriologia Era 3Documento7 páginasAparato Digestivo Embriologia Era 3Milla VieiraAún no hay calificaciones
- Resumen de Sistema Digestivo Embriologia de Langman !!!!Documento6 páginasResumen de Sistema Digestivo Embriologia de Langman !!!!Cedrid MoralesAún no hay calificaciones
- Desarrollo Intestino PrimitivoDocumento74 páginasDesarrollo Intestino PrimitivoSarahi MercadoAún no hay calificaciones
- Desarrollo del tubo digestivoDocumento7 páginasDesarrollo del tubo digestivoCinthya VelazquezAún no hay calificaciones
- Seminario 3 EmbriologíaDocumento32 páginasSeminario 3 EmbriologíaMarco Francesco Barboza AraujoAún no hay calificaciones
- Embriología Del Sistema DigestivoDocumento4 páginasEmbriología Del Sistema DigestivoMilagros NuñezAún no hay calificaciones
- Desarrollo Del Sistema GastrointestinalDocumento56 páginasDesarrollo Del Sistema GastrointestinalNeydi MendezAún no hay calificaciones
- Aparato DigestivoDocumento10 páginasAparato DigestivoJoyce ZambranoAún no hay calificaciones
- Desarrollo de Tubo DigestivoDocumento6 páginasDesarrollo de Tubo Digestivoyviggu100% (1)
- Resumen Embrio 3er ParcialDocumento8 páginasResumen Embrio 3er ParcialFrancisco Emiliani GaliziaAún no hay calificaciones
- Tubo DigestivoDocumento56 páginasTubo DigestivoEmiliano CardenasAún no hay calificaciones
- Aparato Digestivo 2 Embrio 2023Documento4 páginasAparato Digestivo 2 Embrio 2023RaoulAún no hay calificaciones
- Tubo Digestivo - EmbriologíaDocumento22 páginasTubo Digestivo - EmbriologíaOctavio Villacrez OlascoagaAún no hay calificaciones
- Embrio Sistema Digestivo (Resumen)Documento8 páginasEmbrio Sistema Digestivo (Resumen)1er año MED.Aún no hay calificaciones
- Cap 15 EmbrioDocumento14 páginasCap 15 EmbrioBELINDA XCARET RIVERA GONZALEZAún no hay calificaciones
- G.S DIgestivo A)Documento5 páginasG.S DIgestivo A)Adania AguilarAún no hay calificaciones
- Embriologia Del Sistema DigestivoDocumento18 páginasEmbriologia Del Sistema DigestivoDomingo Sanoja GimonAún no hay calificaciones
- Mesos, Celoma y sus anomalíasDocumento5 páginasMesos, Celoma y sus anomalíasAdolfo Antonio Ríos AlcortaAún no hay calificaciones
- 12° Aparato DigestivoDocumento8 páginas12° Aparato DigestivoCarlos RomeroAún no hay calificaciones
- Desarrollo Sistema DigestivoDocumento8 páginasDesarrollo Sistema DigestivoAgustina PerezAún no hay calificaciones
- Sistema DigestivoDocumento42 páginasSistema DigestivoJUAN ENRIQUE CORTEZ QUIROGAAún no hay calificaciones
- Embriologia Del Sistema DigestivoDocumento45 páginasEmbriologia Del Sistema DigestivoJcaminoAún no hay calificaciones
- Capitulo 15 Sistema DigestivoDocumento6 páginasCapitulo 15 Sistema DigestivoRachell CaceresAún no hay calificaciones
- Desarrollo del Aparato DigestivoDocumento6 páginasDesarrollo del Aparato DigestivoZuzeth Marbella García AstorgaAún no hay calificaciones
- Ensayo de CavidadesDocumento12 páginasEnsayo de CavidadesHiyker Xavyer PérEzAún no hay calificaciones
- 10.sistema DigestivoDocumento35 páginas10.sistema DigestivoLuis Alberto Benitez100% (2)
- Tema 12. Intestino Anterior Medio y PosteriorDocumento10 páginasTema 12. Intestino Anterior Medio y PosteriorVibe Nation100% (1)
- Citología ClaseDocumento3 páginasCitología ClasesalazarthisAún no hay calificaciones
- Sistema digestivo embriologíaDocumento6 páginasSistema digestivo embriologíasandra acostaAún no hay calificaciones
- Clase 3 y 4 Embriologia Iii Ciclo 2021 - IiDocumento43 páginasClase 3 y 4 Embriologia Iii Ciclo 2021 - IiTania CAún no hay calificaciones
- 15 DigestivoDocumento18 páginas15 DigestivoSebas YMAún no hay calificaciones
- DesarrolloGIDocumento4 páginasDesarrolloGIMisty LoraAún no hay calificaciones
- Aparato DigestivoDocumento4 páginasAparato DigestivoAlejandro RSAún no hay calificaciones
- Para La Teoria de Morfo El 25Documento3 páginasPara La Teoria de Morfo El 25Taty GueledelAún no hay calificaciones
- Aparato DigestivoDocumento55 páginasAparato DigestivoFernanda CalleAún no hay calificaciones
- Embriología del sistema digestivo. Intestino anterior, medio y posterior en el embrión humanoDocumento54 páginasEmbriología del sistema digestivo. Intestino anterior, medio y posterior en el embrión humanoNAYELI SARAI LOPEZ MELENDEZAún no hay calificaciones
- Aparato Digestivo (TP 8)Documento5 páginasAparato Digestivo (TP 8)Araceli RetamozoAún no hay calificaciones
- Embriología del sistema digestivo enDocumento20 páginasEmbriología del sistema digestivo enCristianAún no hay calificaciones
- CAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaDocumento4 páginasCAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaTodoMePuedeAlvAún no hay calificaciones
- Desarrollo del aparato digestivo humanoDocumento10 páginasDesarrollo del aparato digestivo humanoEduardoAún no hay calificaciones
- DIGESTVO-HistoDocumento65 páginasDIGESTVO-HistoFernanda CalleAún no hay calificaciones
- Intestino Anterior y MedioDocumento6 páginasIntestino Anterior y MedioMarthiitha Carrillo SantiagoAún no hay calificaciones
- Aparato Digestivo Embriologia III ParcialDocumento6 páginasAparato Digestivo Embriologia III ParcialJose PerezAún no hay calificaciones
- Aparato digestivo: desarrollo y formación de órganosDocumento6 páginasAparato digestivo: desarrollo y formación de órganosLeodan Antonio Macas GonzalezAún no hay calificaciones
- Aparato DigestivoDocumento33 páginasAparato DigestivoNICOLE FEENEY CAMACHOAún no hay calificaciones
- RESUMEN KK EMBRIO GASTRODocumento8 páginasRESUMEN KK EMBRIO GASTROliliarroyo0602Aún no hay calificaciones
- Ensayo de CavidadesDocumento11 páginasEnsayo de CavidadesHiyker Xavyer PérEzAún no hay calificaciones
- 15 DigestivoDocumento18 páginas15 DigestivoCarolina Cafezzita PintoAún no hay calificaciones
- Aparato digestivo: divisiones del intestino primitivo y mesenteriosDocumento15 páginasAparato digestivo: divisiones del intestino primitivo y mesenteriosClaudia K50% (2)
- Saco VitelinoDocumento3 páginasSaco VitelinoJACINTO ROSALES YULIZA ZARAIAún no hay calificaciones
- Peritoneo RouviereDocumento9 páginasPeritoneo RouviereMariela Rios100% (1)
- Desarrollo Embriológico Del Aparato DigestivoDocumento2 páginasDesarrollo Embriológico Del Aparato DigestivoRafael María Acacio Jiménez100% (2)
- Fascias de AcolamientoDocumento6 páginasFascias de AcolamientoLore y Juan InglésAún no hay calificaciones
- Sistema DigestDocumento6 páginasSistema DigestSol Arteaga RoaAún no hay calificaciones
- Generalidades Del Sistema Digestiv1Documento46 páginasGeneralidades Del Sistema Digestiv1Ydely RudasAún no hay calificaciones
- Desarrollo de DigestivoDocumento10 páginasDesarrollo de DigestivoAx3L hushAún no hay calificaciones
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- fisiologia digestivaDocumento33 páginasfisiologia digestivaMayary Suarez ZambranoAún no hay calificaciones
- Seminario de AnatomiaDocumento1 páginaSeminario de AnatomiaDrBendezu100% (1)
- Pares Biomagnético Guia CompletaDocumento67 páginasPares Biomagnético Guia CompletaXray UvnAún no hay calificaciones
- Semana 14 EmbrDocumento19 páginasSemana 14 EmbrAnthony CastilloAún no hay calificaciones
- Angelica Viviana Manchinelli OrellanaDocumento78 páginasAngelica Viviana Manchinelli OrellanaysauraAún no hay calificaciones
- Diabetes: Causas, Tipos y ComplicacionesDocumento23 páginasDiabetes: Causas, Tipos y ComplicacionesJeremy UchihaAún no hay calificaciones
- Temario Gastroenterologia 2018Documento2 páginasTemario Gastroenterologia 2018yolanda3nu3ez-645909Aún no hay calificaciones
- Pancreas EmdocrinoDocumento28 páginasPancreas EmdocrinoVladimir ArmijosAún no hay calificaciones
- Anatomia Del Pancreas Rouviere 242046 Downloable 2595396Documento4 páginasAnatomia Del Pancreas Rouviere 242046 Downloable 2595396Juan David HerreraAún no hay calificaciones
- Histologia 3 DR Paz 2020 AlumnoDocumento51 páginasHistologia 3 DR Paz 2020 AlumnoAisha Gordillo HuamanAún no hay calificaciones
- Cuestionario de Biología 3º Eso Sistema Nervioso y EndocrinoDocumento4 páginasCuestionario de Biología 3º Eso Sistema Nervioso y EndocrinoMTHERREROAún no hay calificaciones
- Sindromes Pancreaticos 345348 Downloable 326466Documento7 páginasSindromes Pancreaticos 345348 Downloable 326466jhonatan palominoAún no hay calificaciones
- Sistema neuroendocrino: principales glándulas y funcionesDocumento23 páginasSistema neuroendocrino: principales glándulas y funcionesJaviera Paz Romo GalvezAún no hay calificaciones
- PancreatitisDocumento42 páginasPancreatitisCesar RodasAún no hay calificaciones
- Dieta Cambia Tu MetabolismoDocumento9 páginasDieta Cambia Tu MetabolismoCarlota Vera100% (1)
- Eterminación Del Perfil Hepático de Perros Geriátricos Mediante Pruebas Específicas de LaboratorioDocumento66 páginasEterminación Del Perfil Hepático de Perros Geriátricos Mediante Pruebas Específicas de LaboratorioArturo TurizoAún no hay calificaciones
- Libro Curso 2 Quirofano PDFDocumento186 páginasLibro Curso 2 Quirofano PDFAnonymous RhPJSiRsT0% (1)
- Clase 11 - Enzimas ReguladorasDocumento81 páginasClase 11 - Enzimas ReguladorasJaime Saavedra Guevara100% (3)
- MARIN Marta - Como Dar Fluidez y Cohesion Al TextoDocumento17 páginasMARIN Marta - Como Dar Fluidez y Cohesion Al TextoNatalia AlvarezAún no hay calificaciones
- Glaser, Brigitte y Vogt, Ursula - Reiki (El Poder Sanador de Las Manos)Documento37 páginasGlaser, Brigitte y Vogt, Ursula - Reiki (El Poder Sanador de Las Manos)Belu LucilaAún no hay calificaciones
- Plan de prevención de patologías laborales enfermeríaDocumento10 páginasPlan de prevención de patologías laborales enfermeríaYeraldith CARDENAS HERNANDEZAún no hay calificaciones
- Tumores PancreaticosDocumento44 páginasTumores PancreaticosCarlos Velarde DiazAún no hay calificaciones
- Sistema DigestivoDocumento16 páginasSistema DigestivoManuel Santisteban Tuñoque100% (1)
- Farmacología Del Sistema DigestivoDocumento5 páginasFarmacología Del Sistema DigestivoLuis Daniel Parada100% (1)
- Amilasa PracticaDocumento9 páginasAmilasa PracticaJhenny19950% (1)
- FISIOLOGÍA DEL PÁNCREAS Y SUS FUNCIONES ENDOCRINAS Y EXOCRINASDocumento6 páginasFISIOLOGÍA DEL PÁNCREAS Y SUS FUNCIONES ENDOCRINAS Y EXOCRINASRaúl R. CeledónAún no hay calificaciones
- Tema Endocrino.Documento73 páginasTema Endocrino.Tiitaa OOrozco de TorresAún no hay calificaciones
- Aparato Digestivo en AnimalesDocumento12 páginasAparato Digestivo en Animalestheyears_thehoursAún no hay calificaciones
- Tipos de Tejido EpitelialDocumento7 páginasTipos de Tejido EpitelialMarco RojasAún no hay calificaciones
- PáncreasDocumento14 páginasPáncreasRUBY ALEJANDRA FLOR ANDRADEAún no hay calificaciones