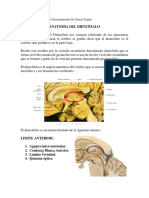
También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- 8 Soluciones Naturales para El Insomnio CronicoDocumento3 páginas8 Soluciones Naturales para El Insomnio CronicoMax MarAún no hay calificaciones
- Cristales Judy Hall Volumen 3Documento160 páginasCristales Judy Hall Volumen 3Andrea Leon100% (2)
- Biomagnetismo Curar Con ImanesDocumento13 páginasBiomagnetismo Curar Con ImanesJuan Jesus Moreno Garcia80% (5)
- Hipotálamo y sistema hipotálamo-hipofisarioDocumento8 páginasHipotálamo y sistema hipotálamo-hipofisarioJose Pablo CopaAún no hay calificaciones
- TalamoDocumento62 páginasTalamoThelma PinayaAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Corteza motora: estructura y funciones principalesDocumento17 páginasCorteza motora: estructura y funciones principalesBryan JuarezAún no hay calificaciones
- Resumen de Neuroanatomía y NeurofisiologíaDocumento8 páginasResumen de Neuroanatomía y NeurofisiologíaRamon CruzAún no hay calificaciones
- El Tálamo y Sus ConexionesDocumento35 páginasEl Tálamo y Sus ConexionesJonathan Ovando67% (3)
- Neurodesarrollo MonografiaDocumento15 páginasNeurodesarrollo MonografiaPs Diego Pérez EgañaAún no hay calificaciones
- El Cerebro y Sus PartesDocumento16 páginasEl Cerebro y Sus PartesOscar Mamani100% (3)
- Manual de La AsignaturaDocumento112 páginasManual de La AsignaturaMaritza Gonzalez Torres Gomez de CadizAún no hay calificaciones
- ForoDocumento16 páginasForoAlexander Aya100% (1)
- Ginecologia y SintergeticaDocumento21 páginasGinecologia y SintergeticaAnonymous oZO05CpaAún no hay calificaciones
- Neuroanatomia - Veterinaria - ContinuacionDocumento5 páginasNeuroanatomia - Veterinaria - ContinuacionMarcela MoraAún no hay calificaciones
- Catedra de Laboratorio de Neuroanatomía Anatomia Del DiencefaloDocumento13 páginasCatedra de Laboratorio de Neuroanatomía Anatomia Del DiencefaloGregory DislaAún no hay calificaciones
- Facultad Ciencias de La Salud Escuela Profesional de OdontologiaDocumento29 páginasFacultad Ciencias de La Salud Escuela Profesional de OdontologiaAndrea SugashimaAún no hay calificaciones
- Areas SubcorticalesDocumento16 páginasAreas SubcorticalesVänëssa ViivancoAún no hay calificaciones
- Erickson Aarón Martínez Tarea 6 HuehueDocumento10 páginasErickson Aarón Martínez Tarea 6 HuehueAndrea VillatoroAún no hay calificaciones
- Trabajo Final de AnatomiaDocumento12 páginasTrabajo Final de AnatomiaDilenia Merejildo100% (1)
- DiencefaloDocumento6 páginasDiencefaloAnais Velasquez MuñozAún no hay calificaciones
- Configuracion Interna e Irrigacion Del Encefalo. OrgDocumento55 páginasConfiguracion Interna e Irrigacion Del Encefalo. OrgMaria Eugenia MendozaAún no hay calificaciones
- Tálamo e HipotalamoDocumento7 páginasTálamo e HipotalamoAnonymous ffFJ7Fngi100% (1)
- Apuntes de Angel Tavera 2do ParcialDocumento14 páginasApuntes de Angel Tavera 2do Parcialjuan peres peresAún no hay calificaciones
- Diencéfalo y Telencéfalo 2013Documento16 páginasDiencéfalo y Telencéfalo 2013Felipe Andres MoyaAún no hay calificaciones
- FB1 2PPDocumento80 páginasFB1 2PPRekkoAún no hay calificaciones
- Sindromes - Piramidales 02112019Documento22 páginasSindromes - Piramidales 02112019Thiago SantosAún no hay calificaciones
- Sistema NerviosoDocumento13 páginasSistema NerviosoLauren ReyesAún no hay calificaciones
- Sistema endocrino: hipotálamo, hipófisis y glándula tiroidesDocumento17 páginasSistema endocrino: hipotálamo, hipófisis y glándula tiroidesChucho MéndezAún no hay calificaciones
- Estructura Interna Del Hemisferio CerebralDocumento83 páginasEstructura Interna Del Hemisferio CerebraldaniinocomeAún no hay calificaciones
- Morfofisiología Del Sistema NerviosoDocumento10 páginasMorfofisiología Del Sistema NerviosoMiguel DuqueAún no hay calificaciones
- PSICOFISIOLOGÍA I_NEUROANATOMÍA FUNCIONAL_SISTEMA LÍMBICODocumento27 páginasPSICOFISIOLOGÍA I_NEUROANATOMÍA FUNCIONAL_SISTEMA LÍMBICOElisabet CascoAún no hay calificaciones
- Sistema NerviosoDocumento18 páginasSistema NerviosoantonioAún no hay calificaciones
- JOSSELYN QUIMBITA Grupo 4 Neoencéfalo, Encéfalo Composición ExternaDocumento56 páginasJOSSELYN QUIMBITA Grupo 4 Neoencéfalo, Encéfalo Composición ExternaMilena QuimbitaAún no hay calificaciones
- Anatomía del hipotálamo - Centro de control vitalDocumento2 páginasAnatomía del hipotálamo - Centro de control vitalMichelle MosqueraAún no hay calificaciones
- Sistema nerviosoDocumento11 páginasSistema nerviosoandreaynoa123Aún no hay calificaciones
- 55Documento10 páginas55angeldavidcuevas2021Aún no hay calificaciones
- 200 años de independencia del PerúDocumento10 páginas200 años de independencia del PerúNicole MoranAún no hay calificaciones
- P3ner PDFDocumento18 páginasP3ner PDFMax George Samame GomezAún no hay calificaciones
- Resumen Neuroanatomía Tema 10Documento10 páginasResumen Neuroanatomía Tema 10Ever RuedaAún no hay calificaciones
- El diencéfalo: tálamo, hipotálamo y másDocumento9 páginasEl diencéfalo: tálamo, hipotálamo y másJuana domingo izquierdoAún no hay calificaciones
- Corteza Motora: Control MovimientosDocumento22 páginasCorteza Motora: Control MovimientosRicardo Pacheco RiosAún no hay calificaciones
- Fichero Fernanda GuevaraDocumento30 páginasFichero Fernanda GuevaraCarlos MArtinezAún no hay calificaciones
- Talamo y Sus Conexiones1Documento41 páginasTalamo y Sus Conexiones1Evely AndarciaAún no hay calificaciones
- Anatomía Del HipotálamoDocumento8 páginasAnatomía Del HipotálamoAlbaAún no hay calificaciones
- Corteza MotoraDocumento18 páginasCorteza MotoraMan LelAún no hay calificaciones
- Anatomía Del Diencéfalo y La Glándula de La HipófisisDocumento17 páginasAnatomía Del Diencéfalo y La Glándula de La HipófisisLucas PichardoAún no hay calificaciones
- Glandula PinealDocumento7 páginasGlandula PinealEmanuel Di Lorenzo100% (1)
- Lectura de ReforzamientoDocumento4 páginasLectura de ReforzamientoGiannina PradoAún no hay calificaciones
- MorfofisiologiaDocumento27 páginasMorfofisiologiamolinapalenciaAún no hay calificaciones
- Sistema Cardiovascular y Nervioso Embrión PolloDocumento15 páginasSistema Cardiovascular y Nervioso Embrión PolloShannon AlexanderAún no hay calificaciones
- Curso Fisioterapia InfantilDocumento51 páginasCurso Fisioterapia InfantilJoxie Oviedo'Aún no hay calificaciones
- Diencefalo, Telencefalo y Sistema LimbicoDocumento39 páginasDiencefalo, Telencefalo y Sistema Limbicolau villazonAún no hay calificaciones
- MesencéfaloDocumento4 páginasMesencéfaloMemen de RuedaAún no hay calificaciones
- Echeverría Meily U3 GuíaDocumento6 páginasEcheverría Meily U3 GuíaMeily BorjasAún no hay calificaciones
- HIPOTÁLAMODocumento5 páginasHIPOTÁLAMOJORGE OLANO OJEDAAún no hay calificaciones
- FISIOLOGÍADocumento5 páginasFISIOLOGÍAguaiqui2001Aún no hay calificaciones
- Neuroprotección del SNCDocumento60 páginasNeuroprotección del SNCYoelys AndreinaAún no hay calificaciones
- Anatomía Macroscópica y Microscópica Del Sistema NerviosoDocumento15 páginasAnatomía Macroscópica y Microscópica Del Sistema NerviosoZuleima Figueredo100% (3)
- Cápsula interna cerebro funciones componentesDocumento7 páginasCápsula interna cerebro funciones componentesRosario AnaAún no hay calificaciones
- El hipotalamo y sus funciones enDocumento6 páginasEl hipotalamo y sus funciones enJenny MoraAún no hay calificaciones
- Teoria Sistema NeuroendocrinoDocumento9 páginasTeoria Sistema NeuroendocrinoBRENDA ELIZABETH QUISPE QUISPEAún no hay calificaciones
- Neuroanatomía y anatomía topográficaDocumento2 páginasNeuroanatomía y anatomía topográficaCelia SalmeronAún no hay calificaciones
- Diencefalo CompressDocumento14 páginasDiencefalo CompresspatriciaAún no hay calificaciones
- La Mente BailarinaDocumento24 páginasLa Mente BailarinaAZAMERAún no hay calificaciones
- Sistema Nervioso Equipo 3 Original1Documento36 páginasSistema Nervioso Equipo 3 Original1jamerica22Aún no hay calificaciones
- Cédula de NegocioDocumento18 páginasCédula de NegocioHector Francisco Colon GarciaAún no hay calificaciones
- Geografia Como CienciaDocumento4 páginasGeografia Como CienciaHector Francisco Colon GarciaAún no hay calificaciones
- Seguridad eléctrica: causas de accidentes y lesionesDocumento35 páginasSeguridad eléctrica: causas de accidentes y lesionesRrLeo Aacc33% (3)
- HormonasDocumento6 páginasHormonasJ U L I E T 011Aún no hay calificaciones
- Modulación Hormonal Del Aprendizaje y La MemoriaDocumento14 páginasModulación Hormonal Del Aprendizaje y La MemoriaJosé Antonio Fernández100% (4)
- Serotonina y AntiserotoninérgicosDocumento49 páginasSerotonina y AntiserotoninérgicosMonserrath GómezAún no hay calificaciones
- Función de La Glándula PituitariaDocumento2 páginasFunción de La Glándula PituitariaFabiola Andrea50% (4)
- Higiene del sueño adolescentesDocumento25 páginasHigiene del sueño adolescentesChio Cariga MacedoAún no hay calificaciones
- Monografia - Sistema-Endocrino-IndiceDocumento27 páginasMonografia - Sistema-Endocrino-IndiceIrmaluz UE50% (4)
- Cronobiologia Resumen 1 PDFDocumento37 páginasCronobiologia Resumen 1 PDFClarita GuerreroAún no hay calificaciones
- Concepto y Caracteristicas de La Glandula PinealDocumento6 páginasConcepto y Caracteristicas de La Glandula Pinealaubrey100% (1)
- KarenJhuliana AmbuludiGuaman PDFDocumento85 páginasKarenJhuliana AmbuludiGuaman PDFa.adolfozevallosAún no hay calificaciones
- Nuevos avances para curar todo tipo de insomnioDocumento12 páginasNuevos avances para curar todo tipo de insomnioFrancisco Javier Méndez CastilloAún no hay calificaciones
- Metabolismo de La MelatoninaDocumento6 páginasMetabolismo de La MelatoninaDaniel Hernandez100% (1)
- 38 3Documento132 páginas38 3Alexis M-cAún no hay calificaciones
- Remedios Caseros Con Hierbas Naturales en Orden AlfabéticoDocumento18 páginasRemedios Caseros Con Hierbas Naturales en Orden AlfabéticoCarlos Alberto CeballosAún no hay calificaciones
- MELATONINADocumento48 páginasMELATONINASrta ChandrikaAún no hay calificaciones
- Suplemento Alimenticio Que Contiene MelatoninaDocumento1 páginaSuplemento Alimenticio Que Contiene Melatoninarsharom3246705Aún no hay calificaciones
- AnsiolíticosDocumento27 páginasAnsiolíticoslourdes leporeAún no hay calificaciones
- Sop MagnesioDocumento10 páginasSop MagnesioAngie QuillaAún no hay calificaciones
- Psicología Fisiologica PEC Curso 13-14Documento2 páginasPsicología Fisiologica PEC Curso 13-14idoceAún no hay calificaciones
- Analisis de La Pelicula El Lado Bueno de Las CosasDocumento14 páginasAnalisis de La Pelicula El Lado Bueno de Las CosasVictoria M. OrtegaAún no hay calificaciones
- Homeopatia y EmocionesDocumento11 páginasHomeopatia y EmocionesJuana Flores CAún no hay calificaciones
- ESTACIONALIDAD REPRODUCTIVA Yuliana ValdésDocumento11 páginasESTACIONALIDAD REPRODUCTIVA Yuliana ValdésYuliana Valdés AguileraAún no hay calificaciones
- Nuestro Cuerpo, Templo Y Morada Del Espíritu Santo: Libro de Sermones Parte 1Documento40 páginasNuestro Cuerpo, Templo Y Morada Del Espíritu Santo: Libro de Sermones Parte 1Estrellita CAAún no hay calificaciones