También podría gustarte
- Micro Bio MaDocumento6 páginasMicro Bio MaOscar F RojasAún no hay calificaciones
- Técnicas ómicas aplicadas al estudio de la microbiotaDe EverandTécnicas ómicas aplicadas al estudio de la microbiotaAún no hay calificaciones
- Monografia PROBIOTICOSDocumento35 páginasMonografia PROBIOTICOSKatherine MartinaAún no hay calificaciones
- 24 Sobrecrecimiento Bacteriano PDFDocumento10 páginas24 Sobrecrecimiento Bacteriano PDFHugh MusicAún no hay calificaciones
- MicrobiotaDocumento9 páginasMicrobiotaAndrés ToscanoAún no hay calificaciones
- Monografia de FisiopatologiaDocumento17 páginasMonografia de FisiopatologiaFiorela CuevaAún no hay calificaciones
- Microbiota IntestinalDocumento50 páginasMicrobiota Intestinaliran ramosAún no hay calificaciones
- Microbiota y probióticos en veterinariaDe EverandMicrobiota y probióticos en veterinariaAún no hay calificaciones
- Microbioma HumanoDocumento9 páginasMicrobioma HumanoMax Rishie DrumAún no hay calificaciones
- Microbiota y AntibioticosDocumento10 páginasMicrobiota y AntibioticosMara HolaAún no hay calificaciones
- Cuáles Son Las Consecuencias de La Desaparición de La Microbiota HumanaDocumento5 páginasCuáles Son Las Consecuencias de La Desaparición de La Microbiota HumanaTyki MikkAún no hay calificaciones
- Resumen Y Guía De Estudio - 10% Humano: Los Microbios Que Habitan En Su Cuerpo Son La Clave Para La Salud Y La FelicidadDe EverandResumen Y Guía De Estudio - 10% Humano: Los Microbios Que Habitan En Su Cuerpo Son La Clave Para La Salud Y La FelicidadCalificación: 1 de 5 estrellas1/5 (1)
- La Vida Interior PDFDocumento5 páginasLa Vida Interior PDFSal EmbAún no hay calificaciones
- El Microbioma HumanoDocumento5 páginasEl Microbioma HumanoJCJQFBTAún no hay calificaciones
- 39 MicrobiotaDocumento10 páginas39 Microbiotamariacarrasquero468Aún no hay calificaciones
- Microbioma Intestinal - MemDocumento11 páginasMicrobioma Intestinal - Memgale diazAún no hay calificaciones
- 67 666 2 PBDocumento7 páginas67 666 2 PBRaquel Liaño de la FuenteAún no hay calificaciones
- El Microbioma Humano LIMPIO (Con Resumenes)Documento18 páginasEl Microbioma Humano LIMPIO (Con Resumenes)Lisa PinelloAún no hay calificaciones
- Articulo CientificoDocumento3 páginasArticulo CientificoAri GuadalupeeAún no hay calificaciones
- Aprendo en Casa Radio 3ro Sem.16Documento3 páginasAprendo en Casa Radio 3ro Sem.16SoledadArriolaOrósAún no hay calificaciones
- MicrobiotaDocumento14 páginasMicrobiotaJimenez AndreaAún no hay calificaciones
- Tema II Cheng 2020Documento14 páginasTema II Cheng 2020Luis Ángel Rod FloAún no hay calificaciones
- MICROBIOTA Lucy Darleny Idrogo BarbozaDocumento14 páginasMICROBIOTA Lucy Darleny Idrogo BarbozaLucy Idrogo BarbozaAún no hay calificaciones
- La Microbiota y El Microbioma Intestinal Humano PDFDocumento7 páginasLa Microbiota y El Microbioma Intestinal Humano PDFYs Ro100% (1)
- Modulo 1 Microbioma y MicrobiotaDocumento101 páginasModulo 1 Microbioma y MicrobiotaPepa GregoriAún no hay calificaciones
- Relación Del Microbiota Intestinal en Patologías Cutáneas: Relationship of The Intestinal Microbiota in Skin PathologiesDocumento9 páginasRelación Del Microbiota Intestinal en Patologías Cutáneas: Relationship of The Intestinal Microbiota in Skin PathologiesCecilia Moyano VegaAún no hay calificaciones
- Bioética, biomedicina y economías de la vida: Hacia una ética de la corresponsabilidadDe EverandBioética, biomedicina y economías de la vida: Hacia una ética de la corresponsabilidadAún no hay calificaciones
- Una Dieta Vegana o Vegetariana Altera Sustancialmente La Microbiota Fecal Colonica Humana - En.esDocumento8 páginasUna Dieta Vegana o Vegetariana Altera Sustancialmente La Microbiota Fecal Colonica Humana - En.esmaria camilaAún no hay calificaciones
- Ma 03790 01Documento4 páginasMa 03790 01YaRe isaAún no hay calificaciones
- Dieta y Microbiota. Impacto en La SaludDocumento5 páginasDieta y Microbiota. Impacto en La SaludNireAún no hay calificaciones
- Tutorial 4 BM FAMED 2020Documento4 páginasTutorial 4 BM FAMED 2020Javier Sebastián Díaz ArellanoAún no hay calificaciones
- Microorganismos de El EstomagoDocumento12 páginasMicroorganismos de El EstomagoJannette NuñezAún no hay calificaciones
- El Microbioma en Enfermedades AutoinmunesDocumento17 páginasEl Microbioma en Enfermedades AutoinmunesKevin OrtizAún no hay calificaciones
- Somos MicrobiosDocumento2 páginasSomos MicrobiosTamara Denisse Aliaga RomeroAún no hay calificaciones
- Und 1. Conceptos Básicos de Microbiología de AlimentosDocumento48 páginasUnd 1. Conceptos Básicos de Microbiología de AlimentosZanyely de la CruzAún no hay calificaciones
- Genero de Las Bacterias para El Funcionamiento Del CuerpoDocumento14 páginasGenero de Las Bacterias para El Funcionamiento Del CuerpoAna SamaniegoAún no hay calificaciones
- Ficha CyT19avaSemDocumento2 páginasFicha CyT19avaSemEduardo MinayaAún no hay calificaciones
- 1 - MicrofloraDocumento13 páginas1 - MicrofloraDr. corazónAún no hay calificaciones
- ALIMENTOS PROBIOTICOS MonografiaDocumento26 páginasALIMENTOS PROBIOTICOS MonografiaNicole Ximena Solis Handabaka100% (1)
- Somos Un Hibrido Humano IIDocumento8 páginasSomos Un Hibrido Humano IIAdolfoLaRocheAún no hay calificaciones
- Tesis Oficial MicrobiotasDocumento11 páginasTesis Oficial Microbiotasmichelle menendezAún no hay calificaciones
- AtomizacionDocumento7 páginasAtomizacionSandra Milena Rojas BuendiaAún no hay calificaciones
- Modulo 6 Microbiota y EmocionesDocumento6 páginasModulo 6 Microbiota y EmocionesTomas SegretAún no hay calificaciones
- Microbiota Intestinal en La Salud y EnfermedadDocumento9 páginasMicrobiota Intestinal en La Salud y EnfermedadLUCIAMELENDEZSAún no hay calificaciones
- Microbiota Humana Y Su Influencia en La Homeostasis Y La Enfermedad ADocumento15 páginasMicrobiota Humana Y Su Influencia en La Homeostasis Y La Enfermedad AEdwin Aquino ApazaAún no hay calificaciones
- La Flora Bacteriana Del Tracto Digestivo - Gastroenterología y HepatologíaDocumento14 páginasLa Flora Bacteriana Del Tracto Digestivo - Gastroenterología y HepatologíamariaAún no hay calificaciones
- UntitledDocumento7 páginasUntitledIsabella BuitragoAún no hay calificaciones
- Funcion de La Microbiana IntestinalDocumento1 páginaFuncion de La Microbiana IntestinalLuis Jeiko ParadaAún no hay calificaciones
- Microbiota Intestinal Humana y DietaDocumento13 páginasMicrobiota Intestinal Humana y DietaElena Valenzuela RamirezAún no hay calificaciones
- 1 s2.0 S0210570521000583 MainDocumento17 páginas1 s2.0 S0210570521000583 MainDanny CortezAún no hay calificaciones
- FICHA 14 CyT 34 PDFDocumento2 páginasFICHA 14 CyT 34 PDFAK AdAún no hay calificaciones
- Información Sobre La Microbiota IntestinalDocumento7 páginasInformación Sobre La Microbiota IntestinalÁngel De Jesús Gómez AlarcónAún no hay calificaciones
- La Micribiota en La Salud e Las Personas.Documento3 páginasLa Micribiota en La Salud e Las Personas.Carlos Daniel Rios OrtegaAún no hay calificaciones
- MICROBIOTADocumento15 páginasMICROBIOTAnanandrealinaresAún no hay calificaciones
- Manual Microbiota PDFDocumento32 páginasManual Microbiota PDFKevin MunguiaAún no hay calificaciones
- Microbiota y Enfermedad 2020.en - EsDocumento22 páginasMicrobiota y Enfermedad 2020.en - Esneptalizuniga114Aún no hay calificaciones
- Casos y Prospectivas en La Industria AlimentariaDocumento36 páginasCasos y Prospectivas en La Industria AlimentariaSandra P MorelAún no hay calificaciones
- La Quimica Del Origen de La VidaDocumento3 páginasLa Quimica Del Origen de La VidaNicole MayorgaAún no hay calificaciones
- Teórico 5 Métodos de EvaluaciónDocumento16 páginasTeórico 5 Métodos de EvaluaciónVale PonceAún no hay calificaciones
- CafeínaDocumento1 páginaCafeínapatricia roblesAún no hay calificaciones
- Farm. Buenas Practicas de Oficina FarmaceuticaDocumento20 páginasFarm. Buenas Practicas de Oficina FarmaceuticaTuxikito Visdey Mas TlvAún no hay calificaciones
- Caracterización Microbiológica y Fisicoquímica de Aguas Subterráneas de Los Municipios de La Paz y San Diego, Cesar, ColombiaDocumento9 páginasCaracterización Microbiológica y Fisicoquímica de Aguas Subterráneas de Los Municipios de La Paz y San Diego, Cesar, ColombiaYorman Zambrano SilvaAún no hay calificaciones
- Mph563 s7 Formato ProyectoDocumento11 páginasMph563 s7 Formato ProyectoVALENTINA FERNANDA GALLARDO SANZANAAún no hay calificaciones
- Linea de TiempoDocumento4 páginasLinea de Tiempoapi-286770384100% (7)
- Convocatoria2dopremionacionalodontologicoDocumento8 páginasConvocatoria2dopremionacionalodontologicoAlvaro Villegas CisnerosAún no hay calificaciones
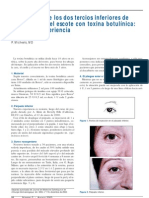
- Articulo BotoxDocumento4 páginasArticulo BotoxViolet Leon Gomez100% (1)
- Agua de HorchataDocumento2 páginasAgua de HorchatajuanAún no hay calificaciones
- Calendario Escolar TecNM Campus PuruandiroDocumento1 páginaCalendario Escolar TecNM Campus PuruandiroCristian AuditoreAún no hay calificaciones
- Ses - Cyt-04-08Documento7 páginasSes - Cyt-04-08Armando Venturo BlasAún no hay calificaciones
- Trauma ObstetricoDocumento18 páginasTrauma ObstetricociclopentatoperidrofAún no hay calificaciones
- Norma Tecnica Ambiental para El Manejo Tratamiento y Disposicion Final de Los Desechos Solidos No-Peligrosos (NTON 05 014-02)Documento31 páginasNorma Tecnica Ambiental para El Manejo Tratamiento y Disposicion Final de Los Desechos Solidos No-Peligrosos (NTON 05 014-02)Jeymi Morelia SuazoAún no hay calificaciones
- INIMPUTABILIDADDocumento20 páginasINIMPUTABILIDADSOCIEDAD DE ALTOS ESTUDIOS DEL DERECHO78% (9)
- Actuación en UrgenciasDocumento7 páginasActuación en Urgenciasjose luisAún no hay calificaciones
- Pauta Guia Sesiones VincularesDocumento7 páginasPauta Guia Sesiones VincularespatriciariquelmevenegasAún no hay calificaciones
- Clinical Pearls in Gastroenterology 2019Documento8 páginasClinical Pearls in Gastroenterology 2019SMIBBtextos textosAún no hay calificaciones
- 011d Hongos Productores de Micosis SubcutáneasDocumento26 páginas011d Hongos Productores de Micosis SubcutáneasJOSELUIS CABREJOS VASQUEZAún no hay calificaciones
- Monografia de Caso ClinicoDocumento15 páginasMonografia de Caso ClinicoDeysi Limaylla NavarroAún no hay calificaciones
- Eduardo Schnitzler El Médico Que Firmó El Parte de Cristina KirchnerDocumento2 páginasEduardo Schnitzler El Médico Que Firmó El Parte de Cristina KirchnerLaNoticia1.comAún no hay calificaciones
- Trabajo de ArtrópodosDocumento5 páginasTrabajo de ArtrópodosAldo Ramón Godoy NuñezAún no hay calificaciones
- Sarval DDocumento4 páginasSarval DAriel De Jesús PerdomoAún no hay calificaciones
- Programa de Educación de La AfectividadDocumento3 páginasPrograma de Educación de La AfectividadJuan Carlos Pérez PérezAún no hay calificaciones
- Vendaje, Inmovilización y TrasladoDocumento36 páginasVendaje, Inmovilización y TrasladoFranco Ibáñez ZumelAún no hay calificaciones
- Lab Rehabilitacion Bucal IIDocumento14 páginasLab Rehabilitacion Bucal IIFernanda GonzalezAún no hay calificaciones
- Practicas Pre - AvanceDocumento11 páginasPracticas Pre - AvanceEfrain CandiottiAún no hay calificaciones
- Precauciones en El Manejo de Material Corto PunzanteDocumento2 páginasPrecauciones en El Manejo de Material Corto PunzanteSergio LLevamealcircoAún no hay calificaciones
- La Cadena EpidemiologicaDocumento6 páginasLa Cadena EpidemiologicaVanesa CaicedoAún no hay calificaciones
- Formato PAE BRAYAN ALVERNIADocumento2 páginasFormato PAE BRAYAN ALVERNIAnatalia bernalAún no hay calificaciones
- Proyecto Capitulo III Kevin Lopez, Jessica SolorzanoDocumento91 páginasProyecto Capitulo III Kevin Lopez, Jessica SolorzanoJULCERISH 1988Aún no hay calificaciones