También podría gustarte
- Guía práctica para técnico superior de laboratorio de diagnóstico clínico y biomédicoDe EverandGuía práctica para técnico superior de laboratorio de diagnóstico clínico y biomédicoCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Nova7 14 24Documento11 páginasNova7 14 24José Luis Cam0% (1)
- KefirDocumento5 páginasKefirMirian Masgo HigidioAún no hay calificaciones
- Recuperación de Levaduras CervecerasDocumento33 páginasRecuperación de Levaduras Cervecerascarlin1905Aún no hay calificaciones
- GUIA El Rol de La Calidad de La Leche en Produccion UHT PDFDocumento43 páginasGUIA El Rol de La Calidad de La Leche en Produccion UHT PDFJuan Pablo AlbarracinAún no hay calificaciones
- Trabajo PPS - Pacheco, Camila - FINALDocumento13 páginasTrabajo PPS - Pacheco, Camila - FINALCamila PachecoAún no hay calificaciones
- Fermentacion Alcoholica de La Pomarrosa-Grupo 5Documento27 páginasFermentacion Alcoholica de La Pomarrosa-Grupo 5Brandon RodriguezAún no hay calificaciones
- Fermentación AlcohólicaDocumento27 páginasFermentación AlcohólicaJocelyn CristernaAún no hay calificaciones
- Sandra Milena Ospina HenaoDocumento83 páginasSandra Milena Ospina HenaoEdson Emmanuel Aldana VásquezAún no hay calificaciones
- Informe VinoDocumento9 páginasInforme VinoMariaAún no hay calificaciones
- Guia TP Micro 2012Documento87 páginasGuia TP Micro 2012roxanachavezpillacaAún no hay calificaciones
- Diseño de Un Plan Haccp para Frijol Molido Enlatado Producido en Alimer S.A. Considerando Evidencia MicrobiológicaDocumento85 páginasDiseño de Un Plan Haccp para Frijol Molido Enlatado Producido en Alimer S.A. Considerando Evidencia MicrobiológicaMax UragaAún no hay calificaciones
- Inmovilizacion de CelulasDocumento10 páginasInmovilizacion de CelulasJimenita CoparaAún no hay calificaciones
- Marco Teórico: Aerobios MesofilosDocumento9 páginasMarco Teórico: Aerobios MesofilosmarlonAún no hay calificaciones
- CAMOTEDocumento147 páginasCAMOTEXhamhuel J D PalominoAún no hay calificaciones
- Aplicaciones de La Ingeniería Genética y La Genómica en EnologíaDocumento9 páginasAplicaciones de La Ingeniería Genética y La Genómica en EnologíaanaiAún no hay calificaciones
- Producción de Ácido Acético Por Erwinia Carotovora A Partir de Zanahoria en Estado de DescomposiciónDocumento19 páginasProducción de Ácido Acético Por Erwinia Carotovora A Partir de Zanahoria en Estado de DescomposiciónVictor TorrecillasAún no hay calificaciones
- Procedimiento Produccion de Puc-Grupo ###2Documento2 páginasProcedimiento Produccion de Puc-Grupo ###2Maira AlejandraAún no hay calificaciones
- .............Documento297 páginas.............jessicaAún no hay calificaciones
- Aislamiento y CaracterizaciónDocumento122 páginasAislamiento y CaracterizaciónChrisAún no hay calificaciones
- Laboratorio 6. BACTERIAS PSICROTRÓPICAS EN LA LECHE PASTEURIZADA (HTST)Documento6 páginasLaboratorio 6. BACTERIAS PSICROTRÓPICAS EN LA LECHE PASTEURIZADA (HTST)Katibel GraellAún no hay calificaciones
- Práctica de Laboratorio para El Aislamiento de MicroorganismosDocumento6 páginasPráctica de Laboratorio para El Aislamiento de MicroorganismosJuan Pablo UrreaAún no hay calificaciones
- FV 29282Documento53 páginasFV 29282LuTentenAún no hay calificaciones
- Taller Conservas Tgcca 2141153Documento8 páginasTaller Conservas Tgcca 2141153D'iana M'artinezAún no hay calificaciones
- Curva de Crecimiento de LevadurasDocumento9 páginasCurva de Crecimiento de LevadurasTavito Acs100% (1)
- Comparación Del Efecto de La Yema de Huevo de Pata y de Gallina SDocumento43 páginasComparación Del Efecto de La Yema de Huevo de Pata y de Gallina SjuraguimaAún no hay calificaciones
- Determinación de La Calidad Microbiológica de Dos Variedades de Quesos, Comercializados en La Zona Uno Del Área Metropolitana de San San SalvadorDocumento110 páginasDeterminación de La Calidad Microbiológica de Dos Variedades de Quesos, Comercializados en La Zona Uno Del Área Metropolitana de San San SalvadorFrancisco HernandezAún no hay calificaciones
- Tesis Paté de Gonadas de PotaDocumento28 páginasTesis Paté de Gonadas de PotaEdgardo QuindeAún no hay calificaciones
- "Técnicas Microbiológicas para El Análisis de La Cerveza": Trabajo Fin de MasterDocumento12 páginas"Técnicas Microbiológicas para El Análisis de La Cerveza": Trabajo Fin de MasterFernando LedesmaAún no hay calificaciones
- Informe 5. Analisis Microbiologico de AlimentosDocumento7 páginasInforme 5. Analisis Microbiologico de AlimentosLUISA BERTHA ADUVIRI CORDOVAAún no hay calificaciones
- Plan Planta LacteosDocumento37 páginasPlan Planta LacteosYuly Petro NegreteAún no hay calificaciones
- Tercer ExamenDocumento10 páginasTercer ExamenVicente MendozaAún no hay calificaciones
- Articulo Cientifico - Carnicos - YamiraDocumento8 páginasArticulo Cientifico - Carnicos - YamiraLizbeth María González CamascaAún no hay calificaciones
- Informe de Fermentacion AlcoholicaDocumento4 páginasInforme de Fermentacion AlcoholicaLess ManriqueAún no hay calificaciones
- Fase 2 Diego Ladino 1022422565Documento11 páginasFase 2 Diego Ladino 1022422565alejandro bossaAún no hay calificaciones
- Yogurt Batido Mediante Sustitución Parcial de Leche Fresca Con Extracto de KiwichaDocumento150 páginasYogurt Batido Mediante Sustitución Parcial de Leche Fresca Con Extracto de KiwichaEdgar Condori Valverde100% (1)
- Guía de Práctica 2Documento3 páginasGuía de Práctica 2MaríaAntezanaAún no hay calificaciones
- Inta Inoculos Innovadores para Enologia Seleccion Hibridos ARTICULODocumento2 páginasInta Inoculos Innovadores para Enologia Seleccion Hibridos ARTICULOMerly Ancco FuentesAún no hay calificaciones
- Lab Micro Recuento de Placa...Documento4 páginasLab Micro Recuento de Placa...Ronny Gamero FernandezAún no hay calificaciones
- Manual Biotecnologia FCienciasDocumento98 páginasManual Biotecnologia FCienciasZak0% (1)
- Trabajos Practicos - Levaduras y Pruebas Bioquimicas - Medina y VirgoliniDocumento14 páginasTrabajos Practicos - Levaduras y Pruebas Bioquimicas - Medina y VirgoliniTisifone95Aún no hay calificaciones
- Copia de Discusión InocuidadDocumento47 páginasCopia de Discusión InocuidadSheyla FuentesAún no hay calificaciones
- Comparacion Levadura de Pan Con Levadura para VinificacionDocumento5 páginasComparacion Levadura de Pan Con Levadura para VinificacionhgyjkllAún no hay calificaciones
- Prácticas Toxiinfecciones AlimentariasDocumento75 páginasPrácticas Toxiinfecciones AlimentariasRobertoAún no hay calificaciones
- Aerobios MesofilosDocumento17 páginasAerobios MesofilosDaniel Luis Solano100% (1)
- Ing. Agroindustrial - Tomas Revilla MendozaDocumento95 páginasIng. Agroindustrial - Tomas Revilla MendozaJulio TaparaAún no hay calificaciones
- Expresion de Amarantina Contra HipertensionDocumento122 páginasExpresion de Amarantina Contra HipertensionHugo Zagal MaldonadoAún no hay calificaciones
- Ensilaje Visceras PescadoDocumento108 páginasEnsilaje Visceras PescadoAQUILINO RAMIREZ SANCHEZ100% (1)
- TFG Maria de La Cruz Santos Navarrete PDFDocumento146 páginasTFG Maria de La Cruz Santos Navarrete PDFFLORENCIAAún no hay calificaciones
- Elaboracion de Vinos 1Documento75 páginasElaboracion de Vinos 1LEONARDO AGUDELOAún no hay calificaciones
- (QO3) Práctica 8 - Síntesis de Biopolímeros (19-19)Documento4 páginas(QO3) Práctica 8 - Síntesis de Biopolímeros (19-19)Daniela A QuezadaAún no hay calificaciones
- Microbiologia IndustrialDocumento77 páginasMicrobiologia IndustrialIvanova Montero SejasAún no hay calificaciones
- LEBAME Estabilidad de Este BioproductoDocumento7 páginasLEBAME Estabilidad de Este BioproductoAlejandroAún no hay calificaciones
- Optimizacin de Variables en Un Biorreactor para La Sntesis de Cido Ctrico Empleando Melaza Como SustratoDocumento32 páginasOptimizacin de Variables en Un Biorreactor para La Sntesis de Cido Ctrico Empleando Melaza Como SustratoPriscilaAún no hay calificaciones
- Trabajo de Bioprospección - Río HirvienteDocumento10 páginasTrabajo de Bioprospección - Río HirvienteNathalieSofia MallquiArgüellesAún no hay calificaciones
- Evaluación Del Potencial Fermentativo de Levaduras Nativas para La Producción de Etanol A Partir de Mieles de Caña de AzúcarDocumento103 páginasEvaluación Del Potencial Fermentativo de Levaduras Nativas para La Producción de Etanol A Partir de Mieles de Caña de AzúcarGian Carlo Salas MendozaAún no hay calificaciones
- Practica MicroDocumento9 páginasPractica MicroSOSA HERNANDEZ YAHAIRA NAOMIAún no hay calificaciones
- Informe de Laboratorio Numero Más ProbableDocumento14 páginasInforme de Laboratorio Numero Más ProbableJulian David Quiroga Sarta60% (5)
- Aislamiento de Cepas de Bacillus Productoras de Proteasas Con Potencial Uso Industrial PDFDocumento68 páginasAislamiento de Cepas de Bacillus Productoras de Proteasas Con Potencial Uso Industrial PDFChristian AndresAún no hay calificaciones
- Factores Que Influyen Sobre La Produccion de La LecheDocumento25 páginasFactores Que Influyen Sobre La Produccion de La LecheTaniaa CaslimAún no hay calificaciones
- 193 22 PBDocumento130 páginas193 22 PByenygirlAún no hay calificaciones
- Acenologa2020 UnqDocumento10 páginasAcenologa2020 UnqyenygirlAún no hay calificaciones
- Natural Occurrence of Ochratoxin A in Musts, Wines and Grape Vine Fruits From Grapes Harvested in Argentina - En.esDocumento13 páginasNatural Occurrence of Ochratoxin A in Musts, Wines and Grape Vine Fruits From Grapes Harvested in Argentina - En.esyenygirlAún no hay calificaciones
- 274 Diciembre 2015 ENCUENTRO VID Botrytis CineDocumento2 páginas274 Diciembre 2015 ENCUENTRO VID Botrytis CineyenygirlAún no hay calificaciones
- 2019 2 Ag050542Documento3 páginas2019 2 Ag050542yenygirlAún no hay calificaciones
- Aminas Biógenas en Alimentos: Métodos Moleculares para La Detección E Identificación de Bacterias ProductorasDocumento15 páginasAminas Biógenas en Alimentos: Métodos Moleculares para La Detección E Identificación de Bacterias ProductorasyenygirlAún no hay calificaciones
- Monográfico: Revista de CienciasDocumento22 páginasMonográfico: Revista de CienciasyenygirlAún no hay calificaciones
- 12 Bioquimica Del VinoDocumento52 páginas12 Bioquimica Del Vinoyenygirl100% (1)
- ¿Qué Son Los Transgénicos U Organismos Genéticamente Modificados (Ogm) ?Documento20 páginas¿Qué Son Los Transgénicos U Organismos Genéticamente Modificados (Ogm) ?jeanevens477Aún no hay calificaciones
- Formulas AntropométricasDocumento58 páginasFormulas AntropométricasErika Iñiguez100% (2)
- Curva de Crecimiento BacterianoDocumento3 páginasCurva de Crecimiento BacterianoDanny Urcia VasquezAún no hay calificaciones
- El Sexo Criminologia OficialDocumento26 páginasEl Sexo Criminologia OficialCesar Chambi ParisacaAún no hay calificaciones
- El Protocolo Bredesen - Es La Nutrición La Clave Del AlzheimerDocumento5 páginasEl Protocolo Bredesen - Es La Nutrición La Clave Del Alzheimerartcelis0% (1)
- Phylum Chytridiomycota 2022Documento47 páginasPhylum Chytridiomycota 2022thepotdrummerallshowAún no hay calificaciones
- Índice: Curso Monitor Mundopark - 2018/19Documento97 páginasÍndice: Curso Monitor Mundopark - 2018/19Laux wnsAún no hay calificaciones
- Guía 2 Reconocimiento de Estructuras FungicasDocumento3 páginasGuía 2 Reconocimiento de Estructuras FungicasBriyid CelennyAún no hay calificaciones
- Mecanismos Biologicos de La TristezaDocumento4 páginasMecanismos Biologicos de La TristezaKatty Ruiz OsorioAún no hay calificaciones
- El Canario BlancoDocumento7 páginasEl Canario BlancoPaco El Zagal ManchegoAún no hay calificaciones
- Pensamiento Critico TaxonomiaDocumento3 páginasPensamiento Critico TaxonomiakarenAún no hay calificaciones
- Bme U1 Ea Lagm 21Documento5 páginasBme U1 Ea Lagm 21Luis Ana100% (1)
- Anammoxosoma PresentacionDocumento49 páginasAnammoxosoma PresentacionPablo VelazquezAún no hay calificaciones
- Exposición Rutas MetabolicasDocumento15 páginasExposición Rutas MetabolicasAlessandro UitzAún no hay calificaciones
- Informe 2 Genética Drosophila MelanogasterDocumento14 páginasInforme 2 Genética Drosophila Melanogasteryuya_solAún no hay calificaciones
- GERMINACIONDocumento17 páginasGERMINACIONG Miller E EspírituAún no hay calificaciones
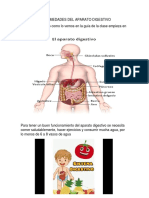
- Enfermedades Del Aparato DigestivoDocumento6 páginasEnfermedades Del Aparato DigestivoKathe Tobon FrancoAún no hay calificaciones
- Seminario Huevo 25879Documento2 páginasSeminario Huevo 25879Christopher VelázquezAún no hay calificaciones
- CoccidiosisDocumento16 páginasCoccidiosisBelen Alvarez GonzalezAún no hay calificaciones
- Bases de Datos de ProteinasDocumento26 páginasBases de Datos de ProteinasAlain VieyraAún no hay calificaciones
- Genética de Las Enfermedades MentalesDocumento3 páginasGenética de Las Enfermedades MentalesMoisés Alejandro Campos Díaz100% (1)
- Genetica Resumen PDFDocumento43 páginasGenetica Resumen PDFSoto Rocío AbigailAún no hay calificaciones
- Los ReplicadoresDocumento24 páginasLos ReplicadoresAlexCancioAún no hay calificaciones
- C U R R I C U L U M V I T A e ClaudiaacevedoDocumento3 páginasC U R R I C U L U M V I T A e ClaudiaacevedoClaudia Acevedo MeinsAún no hay calificaciones
- Guia - 3 Ley de Escala y AlometriaDocumento5 páginasGuia - 3 Ley de Escala y AlometriaAngel David Arrieta MastraAún no hay calificaciones
- Diagnostico Ciencias Naturales 7° BásicoDocumento2 páginasDiagnostico Ciencias Naturales 7° BásicoCindy Iturra Farías85% (95)
- EXAMEN DE BIOLOGÍA ProfDocumento1 páginaEXAMEN DE BIOLOGÍA ProfRamiro Flores MuñozAún no hay calificaciones
- Animales Vertebrados e InvertebradosDocumento19 páginasAnimales Vertebrados e InvertebradosStephanie BenitesAún no hay calificaciones
- 1.fisiopatología de Las InfeccionesDocumento6 páginas1.fisiopatología de Las InfeccionesJAVIERA ROSARIO ZUÑIGA GONZALEZAún no hay calificaciones
- Biología CBC Catedra BanusDocumento4 páginasBiología CBC Catedra BanusjesabelAún no hay calificaciones
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- La increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalDe EverandLa increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalCalificación: 4.5 de 5 estrellas4.5/5 (103)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Historia del cuerpo humano: Evolución, salud y enfermedadDe EverandHistoria del cuerpo humano: Evolución, salud y enfermedadAún no hay calificaciones
- Iniciación al cultivo del cánnabis autoflorecienteDe EverandIniciación al cultivo del cánnabis autoflorecienteCalificación: 5 de 5 estrellas5/5 (3)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Conciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaDe EverandConciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Inteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaDe EverandInteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaAna Isabel Sánchez DíezCalificación: 4 de 5 estrellas4/5 (3)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- PNL: Domina tu mente y aprende como atraer el dinero con técnicas de Programación NeurolingüísticaDe EverandPNL: Domina tu mente y aprende como atraer el dinero con técnicas de Programación NeurolingüísticaCalificación: 4.5 de 5 estrellas4.5/5 (34)
- Diseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaDe EverandDiseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaAún no hay calificaciones
- La biología no miente: Revolución en salud. Basada en las cinco leyes biológicas descubiertas por el Dr. HamerDe EverandLa biología no miente: Revolución en salud. Basada en las cinco leyes biológicas descubiertas por el Dr. HamerCalificación: 4 de 5 estrellas4/5 (4)
- Manual de osteología de cráneo bovino y equinoDe EverandManual de osteología de cráneo bovino y equinoCalificación: 4.5 de 5 estrellas4.5/5 (7)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)