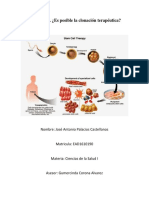
También podría gustarte
- Psicología Animal: Descubre Qué Papel Juega En La VidaDe EverandPsicología Animal: Descubre Qué Papel Juega En La VidaAún no hay calificaciones
- Historia y Conceptos Básicos: Los Orígenes de La Biología Del Desarrollo Un Conjunto de Herramientas ConceptualesDocumento29 páginasHistoria y Conceptos Básicos: Los Orígenes de La Biología Del Desarrollo Un Conjunto de Herramientas ConceptualesGaston LuqueAún no hay calificaciones
- Las células, Constructoras de vida: Cells: Constructing Living ThingsDe EverandLas células, Constructoras de vida: Cells: Constructing Living ThingsCalificación: 3.5 de 5 estrellas3.5/5 (2)
- WWW - Fmed.uba - Ar Depto Histo1a Embrio Adm WolpertDocumento29 páginasWWW - Fmed.uba - Ar Depto Histo1a Embrio Adm Wolpertanon_768966722Aún no hay calificaciones
- Jugos Verdes Le Hablan A Tu Cuerpo - Basado En Las Enseñanzas De Frank Suarez: Descubre Cómo Los Jugos Verdes Impactan Tu CuerpoDe EverandJugos Verdes Le Hablan A Tu Cuerpo - Basado En Las Enseñanzas De Frank Suarez: Descubre Cómo Los Jugos Verdes Impactan Tu CuerpoAún no hay calificaciones
- Wuolah Free Seminario4Documento17 páginasWuolah Free Seminario4Iciar PalomaresAún no hay calificaciones
- 5to Año BiologiaDocumento5 páginas5to Año BiologiaAmadaSAún no hay calificaciones
- 4TA A 8VA SEMANA A PDFDocumento12 páginas4TA A 8VA SEMANA A PDFMaryori Treyci ILasaca ValenzuelaAún no hay calificaciones
- Informe 1Documento11 páginasInforme 1JANET AYME MURILLO VERAAún no hay calificaciones
- 9 DesarrolloembrionarioDocumento12 páginas9 DesarrolloembrionarioAbel PuenteAún no hay calificaciones
- Termino de RegeneracionDocumento7 páginasTermino de RegeneracionツKamixGraphicsAún no hay calificaciones
- Cuadernillo 4toDocumento21 páginasCuadernillo 4toalondrapatronAún no hay calificaciones
- Articulos Diferenciación CelularDocumento7 páginasArticulos Diferenciación CelularGenoveva IbarraAún no hay calificaciones
- DiazMagana Nancy M14S3AI5Documento5 páginasDiazMagana Nancy M14S3AI5Erick DíazAún no hay calificaciones
- Cabrera Aranda - María de Lourdes - M14S3AI5Documento6 páginasCabrera Aranda - María de Lourdes - M14S3AI5Moises Aban CabreraAún no hay calificaciones
- Biologia Del DesarrolloDocumento26 páginasBiologia Del Desarrollomiguerock_90100% (1)
- 10) G4 O. Biologica 2da Parte (Jama)Documento6 páginas10) G4 O. Biologica 2da Parte (Jama)Francis BlackAún no hay calificaciones
- Introducción Al Estudio Del Desarrollo 2017 PDFDocumento66 páginasIntroducción Al Estudio Del Desarrollo 2017 PDFSantiago StrizAún no hay calificaciones
- Filogenia Y Ontogenia Del Sistema NerviosoDocumento5 páginasFilogenia Y Ontogenia Del Sistema NerviosoCamila Garcia OrozcoAún no hay calificaciones
- Semana 1 - Biologia-II-21-32Documento12 páginasSemana 1 - Biologia-II-21-32Jorge ValadzAún no hay calificaciones
- Características de Los Seres VivosDocumento4 páginasCaracterísticas de Los Seres VivosAlfonso Villalobos80% (5)
- Manual EmbriologíaDocumento103 páginasManual EmbriologíaAbel Contreras HerreraAún no hay calificaciones
- Describir El Proceso de Reproducción Asexual en Organismos PluricelularesDocumento18 páginasDescribir El Proceso de Reproducción Asexual en Organismos PluricelularesLipselotte InfanteAún no hay calificaciones
- Informe Biologia 1Documento4 páginasInforme Biologia 1Andres MontesAún no hay calificaciones
- Importancia Del Desarrollo EmbrionarioDocumento2 páginasImportancia Del Desarrollo EmbrionarioSara AGAún no hay calificaciones
- Bio 04 PDFDocumento29 páginasBio 04 PDFyolman26Aún no hay calificaciones
- Articulo Diferenciacion CelularDocumento15 páginasArticulo Diferenciacion CelularYoselyn KisaAún no hay calificaciones
- Documento PDF-DD3C2A93AD10-1Documento8 páginasDocumento PDF-DD3C2A93AD10-1fernanda barragan torresAún no hay calificaciones
- Guía 1 Ciencias Naturales Clei Iii Segundo Periodo BenediktaDocumento9 páginasGuía 1 Ciencias Naturales Clei Iii Segundo Periodo BenediktaJohn Jairo Guzman RoldanAún no hay calificaciones
- Genes animalesDocumento3 páginasGenes animalesMarco A Gonzalez S0% (1)
- Genética de Hongos OriginalDocumento17 páginasGenética de Hongos OriginalGedeon Castillo100% (1)
- Biología Octavo Guía 2. Reproducción Sexual y Asexual 1Documento8 páginasBiología Octavo Guía 2. Reproducción Sexual y Asexual 1YOSELIN ANDREA VILLADIEGO GARCÍAAún no hay calificaciones
- Seres vivos y biomoléculasDocumento21 páginasSeres vivos y biomoléculasAlma HernándezAún no hay calificaciones
- Practica 2Documento32 páginasPractica 2J.L. GaldosAún no hay calificaciones
- S3 Tarea Bases BiologicasDocumento1 páginaS3 Tarea Bases Biologicasvaleria hilarioAún no hay calificaciones
- Informe de Biologia.Documento4 páginasInforme de Biologia.Jade Sikkim Meza MadridAún no hay calificaciones
- Guia de Celulas MadresDocumento10 páginasGuia de Celulas MadresAlvaro Andres Sanchez HurtadoAún no hay calificaciones
- Doc-Orientador (Neurodesarrollo y Educación)Documento12 páginasDoc-Orientador (Neurodesarrollo y Educación)J. Andrea Lemos G.Aún no hay calificaciones
- Formatos y Ejemplos para La Presentación Del Informe de InvestigaciónDocumento17 páginasFormatos y Ejemplos para La Presentación Del Informe de InvestigaciónEzequielAún no hay calificaciones
- BiologiaII 20BDocumento138 páginasBiologiaII 20BIng. Joanna del carmen Anaya AguilarAún no hay calificaciones
- CN08M1_Cómo se replica la vidaDocumento10 páginasCN08M1_Cómo se replica la vidaNorberto MontoyaAún no hay calificaciones
- Guia No. 1 Biologia Grado 6°Documento18 páginasGuia No. 1 Biologia Grado 6°Lozague TammyAún no hay calificaciones
- Sesion 3Documento7 páginasSesion 3emiliano.ir2110Aún no hay calificaciones
- MartinezSalvador Yuliana M14S3Al5Documento4 páginasMartinezSalvador Yuliana M14S3Al5MichelAún no hay calificaciones
- Genética de Hongos - 1Documento10 páginasGenética de Hongos - 1Gedeon CastilloAún no hay calificaciones
- El Origen de La MulticelularidadDocumento8 páginasEl Origen de La MulticelularidadBibiana DuarteAún no hay calificaciones
- Libro AuxologíaDocumento370 páginasLibro Auxologíaanon_287708413Aún no hay calificaciones
- Consulta 6Documento6 páginasConsulta 6Angy PavónAún no hay calificaciones
- Capítulo 4. Meristemos y Diferenciasción de TejidosDocumento23 páginasCapítulo 4. Meristemos y Diferenciasción de Tejidosmyjesa07Aún no hay calificaciones
- The Origin of Metazoa A Unicellular Perspective - En.esDocumento17 páginasThe Origin of Metazoa A Unicellular Perspective - En.esManuel garcia rosasAún no hay calificaciones
- B1 - Texto 3 - Raisman & González 2013 - Características Seres VivosDocumento4 páginasB1 - Texto 3 - Raisman & González 2013 - Características Seres VivosMercy Pacheco PereraAún no hay calificaciones
- Células Troncales y DiferenciaciónDocumento7 páginasCélulas Troncales y DiferenciaciónBlanca TortoledoAún no hay calificaciones
- Actividad 4Documento6 páginasActividad 4PaolaAún no hay calificaciones
- Control Genetico Del Desarrollo EmbrionarioDocumento3 páginasControl Genetico Del Desarrollo EmbrionarioJonathan VzlaAún no hay calificaciones
- Embriología ExperimentalDocumento4 páginasEmbriología Experimentalvalen100% (1)
- Biología SuperiorDocumento6 páginasBiología SuperiorPamela100% (1)
- Actividad 1 Rep. Celular 8a - It 2024Documento4 páginasActividad 1 Rep. Celular 8a - It 2024Diego Armando Acevedo PortillaAún no hay calificaciones
- Evolución y desarrolloDocumento34 páginasEvolución y desarrolloagustinagonzalez525Aún no hay calificaciones
- Características de los seres vivosDocumento31 páginasCaracterísticas de los seres vivosluis robinson bocanegra neyraAún no hay calificaciones
- Intro Ducci OnDocumento2 páginasIntro Ducci OnLiz NohAún no hay calificaciones
- EspermatobiosDocumento11 páginasEspermatobiosRented CribAún no hay calificaciones
- Extracción y Conservación de La Leche Materna.Documento5 páginasExtracción y Conservación de La Leche Materna.Rented CribAún no hay calificaciones
- Contraindicaciones de La Lactancia MaternaDocumento6 páginasContraindicaciones de La Lactancia MaternaRented Crib100% (1)
- Medicamentos y Lactancia Materna.Documento6 páginasMedicamentos y Lactancia Materna.Rented CribAún no hay calificaciones
- Métodos de PriorizaciónDocumento2 páginasMétodos de PriorizaciónRented CribAún no hay calificaciones
- Diagnostico de SaludDocumento1 páginaDiagnostico de SaludRented CribAún no hay calificaciones
- Diseccion RiñonDocumento7 páginasDiseccion RiñonRented CribAún no hay calificaciones
- Importancia de La Adecuada Nutricion Del Recien Nacido y Lactante.Documento8 páginasImportancia de La Adecuada Nutricion Del Recien Nacido y Lactante.Rented CribAún no hay calificaciones
- Composición e inmunología de la leche maternaDocumento12 páginasComposición e inmunología de la leche maternaRented CribAún no hay calificaciones
- Sistema cardiovascular II: Vasos sanguíneosDocumento4 páginasSistema cardiovascular II: Vasos sanguíneosRented CribAún no hay calificaciones
- Semiconductor EsDocumento5 páginasSemiconductor EsRented CribAún no hay calificaciones
- Clonación terapéutica posibleDocumento4 páginasClonación terapéutica posibleGohan Lion Kasamaki OttawaAún no hay calificaciones
- Tipos de ReproducciónDocumento2 páginasTipos de ReproducciónFLORES PALESTINA ALEXANDRA0% (1)
- 3ra Semana de DesarrolloDocumento27 páginas3ra Semana de Desarrollo[FBI] MauroSantanaAún no hay calificaciones
- (PPT 2) Exprecion Genica PDFDocumento7 páginas(PPT 2) Exprecion Genica PDFcamila palominoAún no hay calificaciones
- Cuestionario Sobre Ciclo CelularDocumento4 páginasCuestionario Sobre Ciclo CelularDavid Nieves CamañoAún no hay calificaciones
- Organizador Visual de La Respiracion CelularDocumento1 páginaOrganizador Visual de La Respiracion CelularGabriela Diaz LaureanoAún no hay calificaciones
- Introducción a la genómica grupo 01Documento10 páginasIntroducción a la genómica grupo 01Armando MoraAún no hay calificaciones
- Reproducción Sexual de Las PlantasDocumento8 páginasReproducción Sexual de Las PlantasGiordano ValecillosAún no hay calificaciones
- Planif Refuerzo Pedag CCNN 10 Mo Egb 2023 2024Documento4 páginasPlanif Refuerzo Pedag CCNN 10 Mo Egb 2023 2024ANGEL HERNAN AVILES ZEA100% (1)
- TRANSCRIPCION GeneralidadesDocumento14 páginasTRANSCRIPCION GeneralidadesFacundo Exequiel CorreaAún no hay calificaciones
- TranscripciónDocumento5 páginasTranscripciónDelfi SantellaAún no hay calificaciones
- Procesos de endocitosis, exocitosis y transcitosisDocumento17 páginasProcesos de endocitosis, exocitosis y transcitosisEduardo RenteriaAún no hay calificaciones
- TerpenoidesDocumento12 páginasTerpenoidesCatalina RamirezAún no hay calificaciones
- Taller Individual ReproduccionDocumento2 páginasTaller Individual ReproduccionIDAIRA BARAHONA VERAAún no hay calificaciones
- Preguntas 6 7 8Documento11 páginasPreguntas 6 7 8Yordan Smith BustosAún no hay calificaciones
- Bases de La Reproducción CelularDocumento4 páginasBases de La Reproducción CelularPablo IrustaAún no hay calificaciones
- Aranda Anzaldo Armando La Complejidad y La Forma PDFDocumento91 páginasAranda Anzaldo Armando La Complejidad y La Forma PDFRosalba DuranAún no hay calificaciones
- GENÉTICA DEL SEXO - PPSXDocumento16 páginasGENÉTICA DEL SEXO - PPSXJosé Enrique Domingo OrdóñezAún no hay calificaciones
- ReproducciónDocumento75 páginasReproducciónNADIAAún no hay calificaciones
- Fecundación Y Concepción de Un Ser HumanoDocumento33 páginasFecundación Y Concepción de Un Ser HumanoPesadilla PesadillaAún no hay calificaciones
- Ejercicios Mitosis y Meiosis 1Documento2 páginasEjercicios Mitosis y Meiosis 1Fran IgarzabalAún no hay calificaciones
- Anatomía y Fisiología Del Aparato Reproductor FemeninoDocumento4 páginasAnatomía y Fisiología Del Aparato Reproductor FemeninoGuido Mercado PatiñoAún no hay calificaciones
- Características Reproductivas de La PerraDocumento11 páginasCaracterísticas Reproductivas de La PerraAngela madisonAún no hay calificaciones
- Mapas ConceptualesDocumento12 páginasMapas ConceptualesWarner Rodriguez DuranAún no hay calificaciones
- Estudio de Caso - Division CelularDocumento6 páginasEstudio de Caso - Division CelularCesar CamalAún no hay calificaciones
- Quien Define El Sexo Del BebeDocumento2 páginasQuien Define El Sexo Del BebeBartoloAún no hay calificaciones
- Act 1 y 2 Biologia 2do BGUDocumento2 páginasAct 1 y 2 Biologia 2do BGUMelanie Valeria Quintero PachayAún no hay calificaciones
- Reproducción asexual y sexual en plantasDocumento10 páginasReproducción asexual y sexual en plantasStephany RodríguezAún no hay calificaciones
- Ciclo celular: las etapas de la división celularDocumento1 páginaCiclo celular: las etapas de la división celularEmiliano CastañonAún no hay calificaciones
- Manual Embriología 2018-1Documento25 páginasManual Embriología 2018-1Hugo GonzalezAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- En busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)De EverandEn busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)Calificación: 4.5 de 5 estrellas4.5/5 (3)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Diccionario de neurociencias: Aplicadas a organizaciones y personasDe EverandDiccionario de neurociencias: Aplicadas a organizaciones y personasCalificación: 5 de 5 estrellas5/5 (4)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Plan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolDe EverandPlan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Bases biológicas del comportamiento animal y humanoDe EverandBases biológicas del comportamiento animal y humanoCalificación: 4 de 5 estrellas4/5 (4)
- Mujeres Con Trastornos Por Déficit De Atención: Cómo aceptar sus diferencias y transformar su vidaDe EverandMujeres Con Trastornos Por Déficit De Atención: Cómo aceptar sus diferencias y transformar su vidaCalificación: 3.5 de 5 estrellas3.5/5 (3)
- Especies en comunicación: Aprende a escuchar su vozDe EverandEspecies en comunicación: Aprende a escuchar su vozCalificación: 3.5 de 5 estrellas3.5/5 (7)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Despeja Tu Mente: Como Dejar De Pensar Demasiado, Vencer A Tu Crítico Interno, Y Replantear Tus Pensamientos Negativos Con Hábitos SaludablesDe EverandDespeja Tu Mente: Como Dejar De Pensar Demasiado, Vencer A Tu Crítico Interno, Y Replantear Tus Pensamientos Negativos Con Hábitos SaludablesAún no hay calificaciones
- Hipnoparto Y Parto Creativo: Cómo dar a luz con confianza y sin miedo Altravez del Método LeclaireDe EverandHipnoparto Y Parto Creativo: Cómo dar a luz con confianza y sin miedo Altravez del Método LeclaireCalificación: 4 de 5 estrellas4/5 (7)
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)