También podría gustarte
- Segundos MensajerosDocumento42 páginasSegundos MensajerosMitch Peraza80% (10)
- FISIOLOGÍA DEL SISTEMA ENDOCRINO Páncreas Endocrino Insulin A, Glucagon, ADocumento72 páginasFISIOLOGÍA DEL SISTEMA ENDOCRINO Páncreas Endocrino Insulin A, Glucagon, Aapi-371148392% (13)
- Señalización Celular 2016Documento44 páginasSeñalización Celular 2016Laura Mamani100% (1)
- EnzimologíaDocumento6 páginasEnzimologíaDaniela DiazAún no hay calificaciones
- Sintesis de Insulina PDFDocumento3 páginasSintesis de Insulina PDFXochitemiqui Yoacihualt100% (1)
- Neurotransmisores: Criterios para DenominarlosDocumento28 páginasNeurotransmisores: Criterios para DenominarlosEloíss G. Palacios GallegoAún no hay calificaciones
- NeurotransmisoresDocumento28 páginasNeurotransmisoresAndreaAún no hay calificaciones
- Sustancias Químicas Que Actúan Como Transmisores SinápticosDocumento5 páginasSustancias Químicas Que Actúan Como Transmisores SinápticosPaolaGonzalesAún no hay calificaciones
- Lrpd3-Fundamentos - Briza Arias CasasDocumento17 páginasLrpd3-Fundamentos - Briza Arias CasasBri ACAún no hay calificaciones
- Sind. Hipo e Hipersecreción de InsulinaDocumento38 páginasSind. Hipo e Hipersecreción de Insulinahemer666100% (5)
- Abp TerminadoDocumento8 páginasAbp TerminadoFacundo Mario UmpierrezAún no hay calificaciones
- Taller Fisiología Comunicacion CelularDocumento6 páginasTaller Fisiología Comunicacion CelularLOL X Repleys ElmoshtaykersAún no hay calificaciones
- ENDOCRINO, Anexo Final A Cap Del GuytonDocumento14 páginasENDOCRINO, Anexo Final A Cap Del GuytoncristinaaduartefacultadAún no hay calificaciones
- Guía de Estudios 8. Hormonas - Leyva Vallejo Saul Arturo. Grupo 1Documento9 páginasGuía de Estudios 8. Hormonas - Leyva Vallejo Saul Arturo. Grupo 1arturoleyva2001Aún no hay calificaciones
- Insulina y GlucagondocxDocumento22 páginasInsulina y GlucagondocxTanya Castillo ElizaldeAún no hay calificaciones
- Fisiología General Clase 3Documento35 páginasFisiología General Clase 3diego mendozaAún no hay calificaciones
- Pancreas EndocrinoDocumento8 páginasPancreas EndocrinoAracelis SarmientoAún no hay calificaciones
- Receptores, Segundos Mensajeros y Vias de Senalizacion1Documento14 páginasReceptores, Segundos Mensajeros y Vias de Senalizacion1esrn4laboratorioAún no hay calificaciones
- Receptores, Segundos Mensajeros y Vias de Senalizacion1Documento14 páginasReceptores, Segundos Mensajeros y Vias de Senalizacion1SantiagoAún no hay calificaciones
- Seminario Hormonas UPAO 2018Documento11 páginasSeminario Hormonas UPAO 2018Anonymous vG7Kg83iRAún no hay calificaciones
- Mensajeros IntracelularesDocumento18 páginasMensajeros IntracelularesLeandro RamosAún no hay calificaciones
- Farmacología Del ParasimpáticoDocumento27 páginasFarmacología Del ParasimpáticoMili AlcántaraAún no hay calificaciones
- S03C05 - Membrana (III)Documento42 páginasS03C05 - Membrana (III)Alejandro B. SilvaAún no hay calificaciones
- Hormonas ExpoDocumento24 páginasHormonas Expojosejavier2501Aún no hay calificaciones
- Celular MMMMMDocumento5 páginasCelular MMMMMEmiliana CrisAún no hay calificaciones
- 6 - Teoría - Vereau - Neurotransmisores PDFDocumento37 páginas6 - Teoría - Vereau - Neurotransmisores PDFSol CamusAún no hay calificaciones
- Señalizacion CelularDocumento5 páginasSeñalizacion CelularFaruk CastrilloAún no hay calificaciones
- Mecanismos de Señalización y Acción FarmacológicaDocumento33 páginasMecanismos de Señalización y Acción FarmacológicaOlga Esther Salazar Osuna33% (3)
- DISCUSIONDocumento10 páginasDISCUSIONbryan guidosAún no hay calificaciones
- C2 Farmacodinamia 11-08-22Documento6 páginasC2 Farmacodinamia 11-08-22Sebastian Ignacio Isla PastenAún no hay calificaciones
- Clase 6-Neurotrasmisión Colinérgica - CompressedDocumento25 páginasClase 6-Neurotrasmisión Colinérgica - CompressedMauro Matías CárcamoAún no hay calificaciones
- Vias Señalización Intracitoplasm-1Documento65 páginasVias Señalización Intracitoplasm-1Lizeth Roa RojasAún no hay calificaciones
- INSULINAaDocumento4 páginasINSULINAaGeranni RondonAún no hay calificaciones
- FarmacodinamiaDocumento15 páginasFarmacodinamiamicaabassiAún no hay calificaciones
- Mecanismo de Accion HormonalDocumento5 páginasMecanismo de Accion HormonalPatricio AramburuAún no hay calificaciones
- Fisio-Criterios Parcial 3Documento173 páginasFisio-Criterios Parcial 3Consejo2011Aún no hay calificaciones
- Accion de Los Agentes Neurohumorales Sobre La Membrana y Sobre Los Organos Intracitoplasmaticos Avalos Servin Wendolyn MonserratDocumento4 páginasAccion de Los Agentes Neurohumorales Sobre La Membrana y Sobre Los Organos Intracitoplasmaticos Avalos Servin Wendolyn MonserratVictorJMeloAún no hay calificaciones
- Señalizacion CelularDocumento5 páginasSeñalizacion CelularCIRUGIA MORELOSAún no hay calificaciones
- Fisiologia Del Pancreas y Del Tejido AdiposoDocumento21 páginasFisiologia Del Pancreas y Del Tejido AdipososoniaAún no hay calificaciones
- SEÑALIZACION AutocrinaDocumento20 páginasSEÑALIZACION AutocrinaANAMARIA SANCHEZ TOCORAAún no hay calificaciones
- Adenosín Monofosfato CíclicoDocumento2 páginasAdenosín Monofosfato Cícliconathy00015Aún no hay calificaciones
- Taller PáncreasDocumento15 páginasTaller PáncreasCristian Camilo GonzalezAún no hay calificaciones
- Seminario 3-1Documento7 páginasSeminario 3-1laura sanchez gomisAún no hay calificaciones
- La Transmicion NerviosaDocumento7 páginasLa Transmicion Nerviosaeduardo100% (2)
- ReceptoresDocumento7 páginasReceptoresDennis Russlan Cabrera RochaAún no hay calificaciones
- Taller Del Sistema EndocrinoDocumento13 páginasTaller Del Sistema EndocrinoAurora MuñozAún no hay calificaciones
- Trabajo Escrito. Producción de InsulinaDocumento15 páginasTrabajo Escrito. Producción de InsulinaEdnaAún no hay calificaciones
- Fisio. SIstem. EndocrinoDocumento8 páginasFisio. SIstem. EndocrinoGeranni RondonAún no hay calificaciones
- TP 4 ENDO PancreasDocumento21 páginasTP 4 ENDO PancreasRocio Belen VecchioAún no hay calificaciones
- Tema 2.pancreas Endocrino-1Documento21 páginasTema 2.pancreas Endocrino-1Cordova YolyAún no hay calificaciones
- Examen Final de Fisiologia Animal 2Documento20 páginasExamen Final de Fisiologia Animal 2Jhak AbreguAún no hay calificaciones
- EICOSANOIDESDocumento8 páginasEICOSANOIDESKim ZoldyckAún no hay calificaciones
- Farmacodinamia 1Documento12 páginasFarmacodinamia 1Nahuel RodriguezAún no hay calificaciones
- Tp03 de Bioquímica UNLPDocumento16 páginasTp03 de Bioquímica UNLPAlyson GuilhermeAún no hay calificaciones
- Señales Intracelulares Control GlucosaDocumento13 páginasSeñales Intracelulares Control GlucosaKevin San Martin SanchezAún no hay calificaciones
- Cascadas de SeñalizaciónDocumento10 páginasCascadas de SeñalizaciónCarlos WinchesterAún no hay calificaciones
- Aprenda Farmacología En Una SemanaDe EverandAprenda Farmacología En Una SemanaCalificación: 2 de 5 estrellas2/5 (1)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Prueba de PaternidadDocumento13 páginasPrueba de PaternidadMonroy FernandoAún no hay calificaciones
- Especies de BrucellaDocumento2 páginasEspecies de BrucellaMonroy FernandoAún no hay calificaciones
- NotasDocumento6 páginasNotasMonroy FernandoAún no hay calificaciones
- ShigellaDocumento2 páginasShigellaMonroy FernandoAún no hay calificaciones
- Actualización: Mecanismos Celulares en Respuesta Al Estrés: SirtuinasDocumento12 páginasActualización: Mecanismos Celulares en Respuesta Al Estrés: SirtuinasMonroy FernandoAún no hay calificaciones
- Bordetella PertussisDocumento2 páginasBordetella PertussisMonroy FernandoAún no hay calificaciones
- SalmonellaDocumento4 páginasSalmonellaMonroy Fernando100% (1)
- Clostridium PerfringensDocumento3 páginasClostridium PerfringensMonroy FernandoAún no hay calificaciones
- Clostridium BotulinumDocumento2 páginasClostridium BotulinumMonroy FernandoAún no hay calificaciones
- Intercambio Génico en Los ProcariotasDocumento1 páginaIntercambio Génico en Los ProcariotasMonroy FernandoAún no hay calificaciones
- Adaptaciones Del Crecimiento y La Diferenciación CelularDocumento8 páginasAdaptaciones Del Crecimiento y La Diferenciación CelularMonroy FernandoAún no hay calificaciones
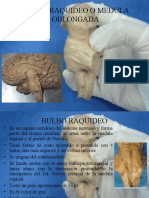
- 1 Bulbo RaquideoDocumento43 páginas1 Bulbo RaquideoMonroy Fernando0% (1)
- Mecanismos de La ApoptosisDocumento7 páginasMecanismos de La ApoptosisMonroy FernandoAún no hay calificaciones
- 2 Puente de VarolioDocumento42 páginas2 Puente de VarolioMonroy FernandoAún no hay calificaciones
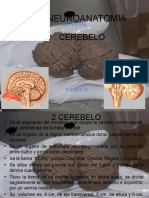
- 3 CerebeloDocumento15 páginas3 CerebeloMonroy FernandoAún no hay calificaciones
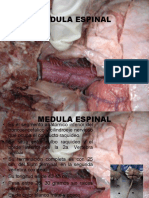
- 5 Medula EspinalDocumento25 páginas5 Medula EspinalMonroy FernandoAún no hay calificaciones
- 4 MesencefaloDocumento20 páginas4 MesencefaloMonroy FernandoAún no hay calificaciones
- Segundos MensajerosDocumento43 páginasSegundos MensajerosMonroy FernandoAún no hay calificaciones
- Sarcoma de Ewing..Documento17 páginasSarcoma de Ewing..Alvaro MartínezAún no hay calificaciones
- 3.esterilización y DesinfecciónDocumento37 páginas3.esterilización y Desinfecciónanthony100% (1)
- ABICDocumento17 páginasABICJaime Martinez HernandezAún no hay calificaciones
- Inmunidad Adaptativa IDocumento44 páginasInmunidad Adaptativa IGustavo Alejandro MolinaAún no hay calificaciones
- Marco Teorico: DefinicionDocumento11 páginasMarco Teorico: DefinicionAriel ZaraviaAún no hay calificaciones
- Dieta para La Helicobacter PyloriDocumento7 páginasDieta para La Helicobacter PyloriMaria Jose Ch100% (1)
- Clase 7 PROTOPLASTO O PROTOPLASMADocumento47 páginasClase 7 PROTOPLASTO O PROTOPLASMAJuan Diego Velez GilAún no hay calificaciones
- Evaluación DietéticaDocumento45 páginasEvaluación DietéticaCris Zamora LunaAún no hay calificaciones
- Alison Sofia CarvajalDocumento11 páginasAlison Sofia CarvajalMaria Deyanira Enciso ArangoAún no hay calificaciones
- 09-06-2003Documento63 páginas09-06-2003La GacetaAún no hay calificaciones
- HepetitisDocumento23 páginasHepetitisOtoniel Cavazos ReynaAún no hay calificaciones
- Accidente Cerebro VascularDocumento8 páginasAccidente Cerebro VascularJesica De SousaAún no hay calificaciones
- Medicamentos Por MembranaDocumento15 páginasMedicamentos Por MembranaMariana NavarroAún no hay calificaciones
- Diagnóstico Diferencial de Las Masas Pulmonares PDFDocumento4 páginasDiagnóstico Diferencial de Las Masas Pulmonares PDFEveryvan DqAún no hay calificaciones
- Dietoterapia Clase 1 Ok.Documento21 páginasDietoterapia Clase 1 Ok.Kristel SolisAún no hay calificaciones
- Cine Foro Resiliencia 2015Documento39 páginasCine Foro Resiliencia 2015clariAún no hay calificaciones
- EVIMALDocumento17 páginasEVIMALAnytta Quilodrán MarquézAún no hay calificaciones
- Genoma HumanoDocumento3 páginasGenoma HumanoYaoh AlejAún no hay calificaciones
- Manual de Procedimiento de Laboratorio para Te PDFDocumento56 páginasManual de Procedimiento de Laboratorio para Te PDFfernie3007Aún no hay calificaciones
- Toxicidad de Los FarmacosDocumento57 páginasToxicidad de Los FarmacosJAJAJAJ100% (2)
- ENZIMAS. Práctica CompletaDocumento10 páginasENZIMAS. Práctica CompletaRafaelAún no hay calificaciones
- Argumentos para El Debate de FolcloreDocumento14 páginasArgumentos para El Debate de FolcloreAlejandro Galicia100% (2)
- Manual 1Documento49 páginasManual 1negritavinanAún no hay calificaciones
- Introducción A La Toxicología AmbientalDocumento35 páginasIntroducción A La Toxicología Ambientalkelly carrizales ramosAún no hay calificaciones
- CEFIXINDocumento4 páginasCEFIXINRachel JohnsonAún no hay calificaciones
- Seminario #04 EpidemioDocumento4 páginasSeminario #04 EpidemiocamilaAún no hay calificaciones
- Interpretación Análisis de SangreDocumento3 páginasInterpretación Análisis de SangreraulgutAún no hay calificaciones
- PeriodoncioDocumento22 páginasPeriodoncioYamaly Mejia100% (1)
- Dialnet AportesDesdeLaPsicologiaAlEstudioDeLaRelacionMente 6269256Documento22 páginasDialnet AportesDesdeLaPsicologiaAlEstudioDeLaRelacionMente 6269256pedro perezAún no hay calificaciones
- Ciclo CelularDocumento14 páginasCiclo CelularEddy Robert Rodriguez BelloAún no hay calificaciones