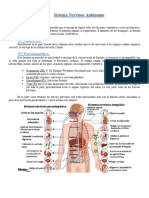
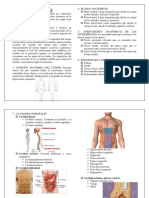
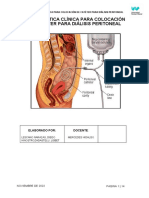
También podría gustarte
- Cap 21 y 22 GujaratiDocumento10 páginasCap 21 y 22 GujaratiErly Apaico Mendoza100% (1)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Trabajo FINAL - Cabrera Eladia BeatrizDocumento18 páginasTrabajo FINAL - Cabrera Eladia BeatrizBeatrizMimyCabreraAún no hay calificaciones
- Nicolás Gómez Dávila. Textos IDocumento75 páginasNicolás Gómez Dávila. Textos Imanuelmorillorubio100% (2)
- Informe Seminario 1Documento19 páginasInforme Seminario 1Mario Santillán RojasAún no hay calificaciones
- Análisis del plan de estudios 2022 de educación básica en MéxicoDocumento112 páginasAnálisis del plan de estudios 2022 de educación básica en MéxicoKarla Patricia Quiroz Aguilar0% (1)
- Introducción al sistema nervioso central (SNC): organización, neurotransmisores y patrones neuronalesDocumento6 páginasIntroducción al sistema nervioso central (SNC): organización, neurotransmisores y patrones neuronalesUzziel LyonAún no hay calificaciones
- Química de La ConductaDocumento10 páginasQuímica de La Conductamanue3lAún no hay calificaciones
- Mecanismos neuronales en los niveles molecular y celularDocumento26 páginasMecanismos neuronales en los niveles molecular y celularZuleik Sanchez Acosta0% (1)
- Neurotransmisores y Neuromoduladores-1Documento26 páginasNeurotransmisores y Neuromoduladores-1Javier Carpio SandovalAún no hay calificaciones
- Neurotransmisores y neuropéptidosDocumento45 páginasNeurotransmisores y neuropéptidosVanessa AvilaAún no hay calificaciones
- Trabajo Síntesis de NeurotrasmisoresDocumento40 páginasTrabajo Síntesis de Neurotrasmisoresfanny monsalveAún no hay calificaciones
- Neuroplasticidad 1Documento11 páginasNeuroplasticidad 1denis rafael noriega espinozaAún no hay calificaciones
- Tejido NerviosoDocumento63 páginasTejido NerviosoyorgeAún no hay calificaciones
- Grupo 2 NEUROBIOLOGÍADocumento7 páginasGrupo 2 NEUROBIOLOGÍAERICK ARISTIDES MENDEZ CABRERAAún no hay calificaciones
- Informe NeuroDocumento15 páginasInforme NeuroDiegolml13Aún no hay calificaciones
- Unidad IiDocumento11 páginasUnidad IiCARLAAún no hay calificaciones
- Neuro Transmi SoresDocumento9 páginasNeuro Transmi Soreshiyuikhnuj100% (1)
- Neurotransmisores Fisiologia Animal.Documento16 páginasNeurotransmisores Fisiologia Animal.Carlos Daniel Quispe ChuraAún no hay calificaciones
- Neurofisiologia (Psicologia)Documento17 páginasNeurofisiologia (Psicologia)Jade JaureguiAún no hay calificaciones
- Seminario 03 CompletoDocumento19 páginasSeminario 03 CompletoReynaldo FaggianiAún no hay calificaciones
- Neuronas y NeurotransmisoresDocumento3 páginasNeuronas y NeurotransmisoresLEVIAún no hay calificaciones
- Apuntes Psicofarmacologia PerseverDocumento50 páginasApuntes Psicofarmacologia PerseverMarina Pizarro SantanoAún no hay calificaciones
- Guía de FarmacologíaDocumento6 páginasGuía de Farmacologíacristian omarAún no hay calificaciones
- Anatomía y Fisiología IIDocumento28 páginasAnatomía y Fisiología IIDaniela Solorzano GonzalezAún no hay calificaciones
- Células glialesDocumento5 páginasCélulas glialesAngelica Tarqui ContrerasAún no hay calificaciones
- NeurotransmisoresDocumento30 páginasNeurotransmisoresAmni JF100% (1)
- CLASE 1 Definición, Descripción y Clasificación de La Neurona y Los Neurotransmisores y Sus ReceptoresDocumento32 páginasCLASE 1 Definición, Descripción y Clasificación de La Neurona y Los Neurotransmisores y Sus ReceptoresMilagrosMoralesZapataAún no hay calificaciones
- Guías Primer Parcial - Andrea Fiamma GarzónDocumento17 páginasGuías Primer Parcial - Andrea Fiamma GarzónFeechu GiustoAún no hay calificaciones
- Sistema Nervioso AutónomoDocumento9 páginasSistema Nervioso AutónomoMonica SaenzAún no hay calificaciones
- CAPITULO 3 - Neurotransmisores y ReceptoresDocumento20 páginasCAPITULO 3 - Neurotransmisores y ReceptoresParaísos PerdidosAún no hay calificaciones
- Arco ReflejoDocumento12 páginasArco ReflejoluikarenAún no hay calificaciones
- 1 NEUROTRANSMISORES 2017 William F. Ganong y Kim E. Barrett - Ganong Fisiología Medica. McGraw-Hill 25va Edición-147-166Documento20 páginas1 NEUROTRANSMISORES 2017 William F. Ganong y Kim E. Barrett - Ganong Fisiología Medica. McGraw-Hill 25va Edición-147-166Eleazar Cadillo Ibañez100% (1)
- Presentacion Cuerpo Humano Organico Ilustrado Morado PastelDocumento43 páginasPresentacion Cuerpo Humano Organico Ilustrado Morado PastelValeria Gómez0% (1)
- Sesión 2 NEURONAS PSICOLOGIADocumento33 páginasSesión 2 NEURONAS PSICOLOGIALuis GalvanAún no hay calificaciones
- Tema 56 6.2 Plantilla The GlobeDocumento14 páginasTema 56 6.2 Plantilla The Globeismael palmaAún no hay calificaciones
- Estructura y función de la neuronaDocumento7 páginasEstructura y función de la neuronaOsvaldo ICAún no hay calificaciones
- Neuronas y NeurotransmisoresDocumento14 páginasNeuronas y NeurotransmisoresEnrique RamosAún no hay calificaciones
- Sem 3 Presencial NEUROTRANSMISORESDocumento27 páginasSem 3 Presencial NEUROTRANSMISORESCarlosAlbertoDangeloToledoAún no hay calificaciones
- Segundo Taller de Fisiologia - NeurotransmisoresDocumento17 páginasSegundo Taller de Fisiologia - NeurotransmisoresJesus manuel bernal petroAún no hay calificaciones
- Botones TerminalesDocumento5 páginasBotones TerminalesBrendita Ramirez0% (1)
- Trabajo Síntesis de NeurotrasmisoresDocumento39 páginasTrabajo Síntesis de Neurotrasmisoresfanny monsalveAún no hay calificaciones
- Neurotransmisores y NeuromoduladoresDocumento21 páginasNeurotransmisores y NeuromoduladoresElizabethEsquivelCruz100% (1)
- NeurotransmisionDocumento11 páginasNeurotransmisionAlberto AmadorAún no hay calificaciones
- Clase de NeurofisioDocumento6 páginasClase de Neurofisiosantiago suarezAún no hay calificaciones
- Introducción A La Farmacología Del SNADocumento36 páginasIntroducción A La Farmacología Del SNAposgradoAún no hay calificaciones
- Sistema nervioso y estudio de la neuronaDocumento15 páginasSistema nervioso y estudio de la neuronaAnthony100% (1)
- NeuropsicologíaDocumento28 páginasNeuropsicologíaCami CabreraAún no hay calificaciones
- Neuronas y Neurotransmisores SinapsisDocumento22 páginasNeuronas y Neurotransmisores SinapsisMARIA YAQUELI LOZANO CHAVEZAún no hay calificaciones
- Moto Neuron ADocumento6 páginasMoto Neuron AFabiola Vásquez CanalesAún no hay calificaciones
- Copia de SNPSDocumento21 páginasCopia de SNPSluccastang9Aún no hay calificaciones
- Seminario 1-Neurotransmisores y NeuropeptidosDocumento25 páginasSeminario 1-Neurotransmisores y NeuropeptidosJimena Custodio GuevaraAún no hay calificaciones
- NEUROTRANSMISORES y NEUROREGULADORES Ok.Documento93 páginasNEUROTRANSMISORES y NEUROREGULADORES Ok.enerAún no hay calificaciones
- AnatonullDocumento24 páginasAnatonullRaysa OrtizAún no hay calificaciones
- Sistema nervioso, sinapsis y neurotransmisoresDocumento5 páginasSistema nervioso, sinapsis y neurotransmisoresangyAún no hay calificaciones
- NEUROTRANSMISORES AGONISTAS-ANTAGONISTASDocumento13 páginasNEUROTRANSMISORES AGONISTAS-ANTAGONISTASAna Ruiz IbáñezAún no hay calificaciones
- NeurotransmisoresDocumento106 páginasNeurotransmisoresJhonnnnnn86% (14)
- NeuronaDocumento42 páginasNeuronaRocio CarrascoAún no hay calificaciones
- Fisiología de la conductaDocumento13 páginasFisiología de la conductamirogomez01Aún no hay calificaciones
- Neurotransmisores y Neuropéptidos - Seminario FisiologíaDocumento32 páginasNeurotransmisores y Neuropéptidos - Seminario FisiologíaJesus Guido Orestes Ojeda TorresAún no hay calificaciones
- Neurotransmisores 40Documento11 páginasNeurotransmisores 40bella esmeralda carbajal mayhuaAún no hay calificaciones
- Sistema Nervioso AutonomoDocumento113 páginasSistema Nervioso AutonomoMARLY EVELYN ROCA CALATAYUDAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- RespuestasDocumento4 páginasRespuestasJoselyne Freire PalmaAún no hay calificaciones
- ClasesDocumento22 páginasClasesJoselyne Freire PalmaAún no hay calificaciones
- Bioquímica del sueñoDocumento5 páginasBioquímica del sueñoJoselyne Freire PalmaAún no hay calificaciones
- Cuadro 2Documento1 páginaCuadro 2Joselyne Freire PalmaAún no hay calificaciones
- Razonamiento Formal y ContenidoDocumento1 páginaRazonamiento Formal y ContenidoJoselyne Freire PalmaAún no hay calificaciones
- 1 Guia Tecnica Planeacion Docentes Preescolar PDFDocumento22 páginas1 Guia Tecnica Planeacion Docentes Preescolar PDFIsis DalíAún no hay calificaciones
- NENOIN Management of Peripheral Neuropathy With B1 B6 and B12 - Jan 2018Documento28 páginasNENOIN Management of Peripheral Neuropathy With B1 B6 and B12 - Jan 2018Angelal100% (1)
- Instrumento de Autoevaluación Tramo Inicial EMERSONDocumento4 páginasInstrumento de Autoevaluación Tramo Inicial EMERSONgracielamerlo2008Aún no hay calificaciones
- Investigación preliminar por presunta usurpación archivarDocumento5 páginasInvestigación preliminar por presunta usurpación archivarVictor ArjonaAún no hay calificaciones
- Ecuaciones Diferenciales Parciales: Clasificación y TiposDocumento2 páginasEcuaciones Diferenciales Parciales: Clasificación y TiposVero ValdezzAún no hay calificaciones
- Act03 Formato de Bitacora COL de Tercer NivelDocumento4 páginasAct03 Formato de Bitacora COL de Tercer NivelRamon EscalanteAún no hay calificaciones
- University en 1959Documento3 páginasUniversity en 1959RECURSOS HUMANOSAún no hay calificaciones
- FICHA El EscepticismoDocumento7 páginasFICHA El EscepticismoCamila LopezAún no hay calificaciones
- Foraminíferos, ostrácodos y calciesferas en formaciones del Paleozoico y Mesozoico de GuatemalaDocumento22 páginasForaminíferos, ostrácodos y calciesferas en formaciones del Paleozoico y Mesozoico de GuatemalaGabriel E Morales RecinosAún no hay calificaciones
- Como Sistematizar en Cinco TiemposDocumento11 páginasComo Sistematizar en Cinco TiemposMILEAún no hay calificaciones
- Laboratorio Bioquímica URLDocumento14 páginasLaboratorio Bioquímica URLHugo CAún no hay calificaciones
- Antologia COI NOI SAEDocumento110 páginasAntologia COI NOI SAE100473367% (3)
- 2020A - Física - 12 - Productos Entre Vectores - Producto Vectorial o Producto Cruz PDFDocumento4 páginas2020A - Física - 12 - Productos Entre Vectores - Producto Vectorial o Producto Cruz PDFjuanAún no hay calificaciones
- UNI-INGENIERÍA ECONÓMICA Y FINANZASDocumento7 páginasUNI-INGENIERÍA ECONÓMICA Y FINANZASHerles Marco Puma PolancoAún no hay calificaciones
- Activity Guide and Evaluation Rubric - Unit 2 - Task 4 - Creating A City - En.esDocumento6 páginasActivity Guide and Evaluation Rubric - Unit 2 - Task 4 - Creating A City - En.esEl jojo's de tu vida100% (1)
- Clase virtual de gerencia sobre trabajo remotoDocumento10 páginasClase virtual de gerencia sobre trabajo remotojose mostaceroAún no hay calificaciones
- ANTIPARKINSONIANOS FinDocumento32 páginasANTIPARKINSONIANOS Finivanpch456823Aún no hay calificaciones
- Primeros Auxilios - AnatomiaDocumento10 páginasPrimeros Auxilios - AnatomiaHAROLD100% (1)
- Caso Práctico Mercadeo EstratégicoDocumento3 páginasCaso Práctico Mercadeo EstratégicoAngel LemusAún no hay calificaciones
- El Pensamiento CentroamericanoDocumento26 páginasEl Pensamiento CentroamericanoEver Fuentes100% (1)
- Cantos Del Vía Crucis GuitarraDocumento6 páginasCantos Del Vía Crucis GuitarrarycerzniepokalanejAún no hay calificaciones
- El Origen Del CorridoDocumento49 páginasEl Origen Del CorridoRicardo Hernandez AguilarAún no hay calificaciones
- Desventajas de Los Costos Basados en Actividades ABCDocumento2 páginasDesventajas de Los Costos Basados en Actividades ABCleticia EchegoyenAún no hay calificaciones
- Verein Deutscher Ingenieure Metodologia de La Investigacion para El DiseñoDocumento1 páginaVerein Deutscher Ingenieure Metodologia de La Investigacion para El DiseñoRicardo Carrillo EsparzaAún no hay calificaciones
- Lenguaje en SytDocumento69 páginasLenguaje en Sytvirgi-sAún no hay calificaciones
- Colocacion CATETER PERITONEALDocumento14 páginasColocacion CATETER PERITONEALLisbet Hinostroza GasteluAún no hay calificaciones