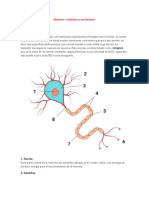
También podría gustarte
- Tejido NerviosoDocumento7 páginasTejido NerviosoAlma HaeAún no hay calificaciones
- Sistema Nervioso CienciasDocumento3 páginasSistema Nervioso CienciasBaezAlvAún no hay calificaciones
- Tejido Nerviso 1, 2do AñoDocumento10 páginasTejido Nerviso 1, 2do AñoMercedes SánchezAún no hay calificaciones
- Fisiologia de La Neurona IDocumento7 páginasFisiologia de La Neurona IJaviera AriasAún no hay calificaciones
- PsicologiaDocumento6 páginasPsicologiamindrycuevas7Aún no hay calificaciones
- Practico No 2 de Fundamentos NeurobiologicosDocumento10 páginasPractico No 2 de Fundamentos Neurobiologicosricky_2014Aún no hay calificaciones
- Tejido Nervioso y Sistema NerviosoDocumento16 páginasTejido Nervioso y Sistema Nerviosomsolorzano-1Aún no hay calificaciones
- UNIDAD 6 Comprimido 2Documento13 páginasUNIDAD 6 Comprimido 2youcanfindme5678Aún no hay calificaciones
- Infotme de Tejido Nervioso y HematopoyeticoDocumento30 páginasInfotme de Tejido Nervioso y HematopoyeticoRotsen MoralesAún no hay calificaciones
- Histo. Semana 5. Teoria. Sistema Nervioso IDocumento50 páginasHisto. Semana 5. Teoria. Sistema Nervioso ILeonardo MarrufoAún no hay calificaciones
- mdml502 s1 Valenzuela1Documento10 páginasmdml502 s1 Valenzuela1Larissa Noemi Saavedra ZuberAún no hay calificaciones
- Estructura y Morfología de La NeuronaDocumento12 páginasEstructura y Morfología de La NeuronaLidyane Aracy Asmat Ulloque100% (1)
- Preguntas de Tejido Nervioso.Documento23 páginasPreguntas de Tejido Nervioso.Arturo ToGa100% (1)
- Apuntes Tejido Nervioso PDFDocumento10 páginasApuntes Tejido Nervioso PDFFernandoAlioAún no hay calificaciones
- Biología 04 5to BachDocumento2 páginasBiología 04 5to BachLibreria BelenAún no hay calificaciones
- Clase 5 Biología Celular de Las NeuronasDocumento74 páginasClase 5 Biología Celular de Las Neuronasjrma003156Aún no hay calificaciones
- Fibras nerviosas y sistema nervioso periféricoDocumento65 páginasFibras nerviosas y sistema nervioso periféricovegalyAún no hay calificaciones
- Histologia Del Tejido NerviosoDocumento48 páginasHistologia Del Tejido NerviosoElias Castillo Castellanos100% (1)
- Identificar Los Componentes Esenciales de La NeuronaDocumento13 páginasIdentificar Los Componentes Esenciales de La NeuronaTam OrtizAún no hay calificaciones
- Partes de Las NeuronasDocumento34 páginasPartes de Las NeuronasRamon Doroteo Mansilla100% (1)
- Fisiologia - Sistema - Nervioso - PPT ExpoDocumento45 páginasFisiologia - Sistema - Nervioso - PPT ExpoSalud Medicina Confianza0% (1)
- Estructura y funciones de la neurona enDocumento3 páginasEstructura y funciones de la neurona ensandra juliana nuñez lambrañoAún no hay calificaciones
- Partes de Una NeuronaDocumento4 páginasPartes de Una NeuronaDiego Alonso Carrera RealpeAún no hay calificaciones
- Tejido NerviosoDocumento12 páginasTejido NerviosoTomás JofreAún no hay calificaciones
- ESTRUCTURA DE LA NEURONADocumento6 páginasESTRUCTURA DE LA NEURONAGabriela Díaz CiezaAún no hay calificaciones
- Tema I Neuronas y Redes Neuronales PDF - CompressedDocumento34 páginasTema I Neuronas y Redes Neuronales PDF - CompressedAndrea valencia vélezAún no hay calificaciones
- Neuron ADocumento5 páginasNeuron Amax_14_15Aún no hay calificaciones
- Sistema Nervioso PisanoDocumento34 páginasSistema Nervioso PisanonicolasAún no hay calificaciones
- La neurona: estructura y clasificacionesDocumento6 páginasLa neurona: estructura y clasificacionesNerea Rodríguez GómezAún no hay calificaciones
- Transmisión Del Impulso NerviosoDocumento35 páginasTransmisión Del Impulso NerviosoGustavo Luis FerriAún no hay calificaciones
- Tejido NerviosoDocumento17 páginasTejido NerviosoMelany MolinaAún no hay calificaciones
- Tejido Nervioso Material ComplementarioDocumento19 páginasTejido Nervioso Material Complementariomiguel bravoAún no hay calificaciones
- taREA nEUROFISIOLOGIADocumento7 páginastaREA nEUROFISIOLOGIAElisa RodríguezAún no hay calificaciones
- La Neurona ExpoDocumento28 páginasLa Neurona ExpoMarilynMolinaAún no hay calificaciones
- Una Neurona Típica Consta deDocumento5 páginasUna Neurona Típica Consta deMargarita Piña RodriguezAún no hay calificaciones
- Tema 6 Organización General Del Sistema NerviosoDocumento52 páginasTema 6 Organización General Del Sistema NerviosoGaia OrevAún no hay calificaciones
- Resumen de Histología de La Medula EspinalDocumento8 páginasResumen de Histología de La Medula EspinalLucia Saavedra LozadaAún no hay calificaciones
- Tema 6. Organizacion General Del Sistema NerviosoDocumento52 páginasTema 6. Organizacion General Del Sistema NerviosoCarlos Javier Nogales Fuentes100% (1)
- Morán García Yairt Hait 1IV6Documento2 páginasMorán García Yairt Hait 1IV6Morán García Yairt HaitAún no hay calificaciones
- Neuronas y su función en el sistema nerviosoDocumento14 páginasNeuronas y su función en el sistema nerviosoLiramy Pierinna ReyesAún no hay calificaciones
- INFORMEDocumento27 páginasINFORMEAshly FloresAún no hay calificaciones
- Fisiologia Sensorial de La Via OpticaDocumento45 páginasFisiologia Sensorial de La Via OpticaGabrielaAún no hay calificaciones
- NeuronaDocumento8 páginasNeuronaReyna RodriguezAún no hay calificaciones
- La NeuronaDocumento11 páginasLa NeuronaHenry W. Montesinos LimaAún no hay calificaciones
- Apuntes Tejido Nervioso PDFDocumento17 páginasApuntes Tejido Nervioso PDFDiego Rosso JaraAún no hay calificaciones
- Primer ParcialDocumento95 páginasPrimer ParcialVale IsbarboAún no hay calificaciones
- Resumen NeuroDocumento137 páginasResumen NeuroMauricio VianaAún no hay calificaciones
- La NeuronaDocumento17 páginasLa NeuronaAlejandra CoronadoAún no hay calificaciones
- Tejido Nervioso y Sus Características Del TejidoDocumento4 páginasTejido Nervioso y Sus Características Del TejidoIssei SenpaiAún no hay calificaciones
- Bases N Trab #4Documento8 páginasBases N Trab #4Caro GeAún no hay calificaciones
- Introducción A La Neurociencia - BrainwareDocumento51 páginasIntroducción A La Neurociencia - Brainwaremateo pianaAún no hay calificaciones
- Resumen Tejido NerviosoDocumento14 páginasResumen Tejido NerviosoMaria Coto Racero100% (4)
- Psicobiología Tema 6 ResumenDocumento10 páginasPsicobiología Tema 6 ResumenArabiaDíazAún no hay calificaciones
- Neurona - WikipediaDocumento7 páginasNeurona - WikipediaVíctor MamaniAún no hay calificaciones
- Introduccion Basica VV 0.202110 PDFDocumento19 páginasIntroduccion Basica VV 0.202110 PDFJeannette del RosarioAún no hay calificaciones
- Apuntes de Neuroanatomia UAH 2014Documento214 páginasApuntes de Neuroanatomia UAH 2014Dayana100% (2)
- Unidad 3Documento18 páginasUnidad 3Leonel NoyaAún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Mapa Conceptual Bases Biologica de La Conducta 2Documento1 páginaMapa Conceptual Bases Biologica de La Conducta 2jonathanca1086% (22)
- Mapa Mental 10Documento6 páginasMapa Mental 10chapis885Aún no hay calificaciones
- Emociones y resiliencia en familiaDocumento5 páginasEmociones y resiliencia en familiaGuadalupeAún no hay calificaciones
- La Neurona: Lic. César Navarro RetuertoDocumento21 páginasLa Neurona: Lic. César Navarro RetuertoGuadalupeAún no hay calificaciones
- Construye Un Mapa Conceptual de Los Tipos de Neuronas Explicando Sus Funciones.Documento1 páginaConstruye Un Mapa Conceptual de Los Tipos de Neuronas Explicando Sus Funciones.GuadalupeAún no hay calificaciones
- METASDocumento2 páginasMETASGuadalupeAún no hay calificaciones
- Ingles Recuperación.2 Trimestre.Documento6 páginasIngles Recuperación.2 Trimestre.GuadalupeAún no hay calificaciones
- Mantenimiento de herramientas, máquinas e instrumentosDocumento5 páginasMantenimiento de herramientas, máquinas e instrumentosGuadalupeAún no hay calificaciones
- EDUC.F. 1o Sem.33,34 NUEVADocumento4 páginasEDUC.F. 1o Sem.33,34 NUEVAGuadalupeAún no hay calificaciones
- Crea diseños de moda a partir de atmósferas y sensopercepcionesDocumento3 páginasCrea diseños de moda a partir de atmósferas y sensopercepcionesGuadalupeAún no hay calificaciones
- Ficha 8. - 13 - Español - 1°Documento5 páginasFicha 8. - 13 - Español - 1°GuadalupeAún no hay calificaciones
- Futbol.: Educaci Ón FísicaDocumento4 páginasFutbol.: Educaci Ón FísicaGuadalupeAún no hay calificaciones
- VIDA PANDEMIADocumento2 páginasVIDA PANDEMIAGuadalupeAún no hay calificaciones
- FICHA 38 y 39 ARTES SECUNDARIA 1o OficialDocumento4 páginasFICHA 38 y 39 ARTES SECUNDARIA 1o OficialGuadalupeAún no hay calificaciones
- Grado Ficha 25 y 26 Fce IDocumento2 páginasGrado Ficha 25 y 26 Fce IGuadalupeAún no hay calificaciones
- JunioDocumento5 páginasJunioGuadalupeAún no hay calificaciones
- FICHA 29, 30 y 31 NUEVADocumento3 páginasFICHA 29, 30 y 31 NUEVAGuadalupeAún no hay calificaciones
- Ficha 29y30 NuevaDocumento3 páginasFicha 29y30 NuevaGuadalupe100% (1)
- Semana 29-30-31 NuevaDocumento4 páginasSemana 29-30-31 NuevaGuadalupeAún no hay calificaciones
- Ficha 36 J37 de GeografiaDocumento6 páginasFicha 36 J37 de GeografiaGuadalupeAún no hay calificaciones
- Ficha 29y30 NuevaDocumento3 páginasFicha 29y30 NuevaGuadalupe100% (1)
- NUEVADocumento7 páginasNUEVAGuadalupeAún no hay calificaciones
- NUEVADocumento4 páginasNUEVAGuadalupeAún no hay calificaciones
- Mantenimiento de herramientas, máquinas e instrumentosDocumento5 páginasMantenimiento de herramientas, máquinas e instrumentosGuadalupeAún no hay calificaciones
- Ficha 27, 28de Geografia FICHA NUEVADocumento3 páginasFicha 27, 28de Geografia FICHA NUEVAGuadalupeAún no hay calificaciones
- Ficha Socioemocional 1,2y3 1 Al 12 MarzoDocumento4 páginasFicha Socioemocional 1,2y3 1 Al 12 MarzoGuadalupeAún no hay calificaciones
- Inglés 1.sem.27-28 Ficha NuevaDocumento9 páginasInglés 1.sem.27-28 Ficha NuevaGuadalupeAún no hay calificaciones
- Inglés 1 - Sem.25-26Documento6 páginasInglés 1 - Sem.25-26GuadalupeAún no hay calificaciones
- Inglés 1 - Sem.25-26Documento6 páginasInglés 1 - Sem.25-26GuadalupeAún no hay calificaciones
- Clase N°3 Crecimiento CraneofacialDocumento52 páginasClase N°3 Crecimiento CraneofacialSandy Hernández MalcaAún no hay calificaciones
- URG - Articulaciones de La CabezaDocumento38 páginasURG - Articulaciones de La CabezaXimena Olivares CastilloAún no hay calificaciones
- Citogine (1) .OdtDocumento48 páginasCitogine (1) .OdtMeli BashAún no hay calificaciones
- Práctica 1 GastroDocumento12 páginasPráctica 1 GastroAndrea RosasAún no hay calificaciones
- Corazón HumanoDocumento6 páginasCorazón HumanoRichard Anthony Sanchez PonteAún no hay calificaciones
- Biologia Del Crecimiento TumoralDocumento28 páginasBiologia Del Crecimiento Tumoralrachel penuelaAún no hay calificaciones
- Sistemas interconectadosDocumento4 páginasSistemas interconectadosIsa ChaparroAún no hay calificaciones
- Eje Hipotalamo Hipofisis GonadalDocumento8 páginasEje Hipotalamo Hipofisis GonadalIsra BaronaAún no hay calificaciones
- Sistema CardiovascularDocumento200 páginasSistema CardiovascularJuan David Arias ForeroAún no hay calificaciones
- Generalidades de la artrologíaDocumento17 páginasGeneralidades de la artrologíakaalipsooAún no hay calificaciones
- Composicion Del Plasma SanguineoDocumento3 páginasComposicion Del Plasma SanguineoManubri100% (1)
- Pdfsrmacma 2015cma154i PDFDocumento8 páginasPdfsrmacma 2015cma154i PDFAngélica ContrerasAún no hay calificaciones
- Atlas de Anatomía ProyectoDocumento98 páginasAtlas de Anatomía ProyectoIvan Esteban Carreño Calderon100% (2)
- Atención Enfermera en El Paciente Con Alteraciones RespiratoriasDocumento224 páginasAtención Enfermera en El Paciente Con Alteraciones Respiratoriaspastizal123456Aún no hay calificaciones
- Mecánica de La Ventilación PulmonarDocumento56 páginasMecánica de La Ventilación PulmonaromarAún no hay calificaciones
- Desarrollo facial y estructuras oralesDocumento59 páginasDesarrollo facial y estructuras oralesBrandon ChavezAún no hay calificaciones
- Introducción A La Anatomía PatológicaDocumento50 páginasIntroducción A La Anatomía PatológicaAlexandry Ocas Jara100% (2)
- Departamento de Ciencias Morfologicas Componente Histologia Humana IiiDocumento3 páginasDepartamento de Ciencias Morfologicas Componente Histologia Humana IiiDervin Dariel Sevilla OlivasAún no hay calificaciones
- Célula HumanaDocumento6 páginasCélula HumanaLeslie ZarateAún no hay calificaciones
- 5 PDFDocumento29 páginas5 PDFJuan David SanchezAún no hay calificaciones
- Fisiología Respiratoria - Edic West 10a EdDocumento240 páginasFisiología Respiratoria - Edic West 10a Edada100% (1)
- Pparte 3Documento3 páginasPparte 3Dennys OrozcoAún no hay calificaciones
- Informe ErgometríaDocumento5 páginasInforme ErgometríaJuan marcelo CorsichAún no hay calificaciones
- NeuroDocumento10 páginasNeuroOliver Moreno MartinezAún no hay calificaciones
- Biologia Trabajo CelulaDocumento32 páginasBiologia Trabajo CelulaKARLA BAUTISTAAún no hay calificaciones
- Guía de Práctica de HistologíaDocumento79 páginasGuía de Práctica de HistologíaPiero Eduardo Ponce Lingán100% (1)
- Hemograma 11.10Documento6 páginasHemograma 11.10luz marina sosa condoriAún no hay calificaciones
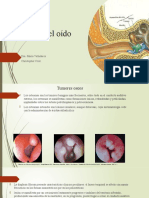
- Tumores Del Oído: Dra. Maria Valladares Christopher CruzDocumento12 páginasTumores Del Oído: Dra. Maria Valladares Christopher CruzCHRISTOPHER SAHIR CRUZ COLINDRESAún no hay calificaciones
- Ciclo MenstrualDocumento49 páginasCiclo MenstrualElena AlvaradoAún no hay calificaciones
- Guia de Aprendizaje La Circulacion 5º Julio Del 2020Documento14 páginasGuia de Aprendizaje La Circulacion 5º Julio Del 2020Lizbeth marinAún no hay calificaciones