También podría gustarte
- ELECTROENCEFALOGRAMADocumento23 páginasELECTROENCEFALOGRAMAJulio VázquezAún no hay calificaciones
- Bases Celulares EEG PlomoDocumento26 páginasBases Celulares EEG PlomoJefferson Collachagua DelgadoAún no hay calificaciones
- Mapeo Cerebral y Enfermedad PDFDocumento30 páginasMapeo Cerebral y Enfermedad PDFRodolfo Gómez Sánchez Aguilar100% (1)
- Pruebas de Evaluación de Funciones CorporalesDocumento73 páginasPruebas de Evaluación de Funciones CorporalesJuniorAún no hay calificaciones
- EEG - ClasesDocumento33 páginasEEG - ClasesMarcos GiorgioAún no hay calificaciones
- 2.1 Tema-EncefalogramaDocumento43 páginas2.1 Tema-Encefalogramacarlota100% (1)
- Sistema de Medición de Señales EEGDocumento8 páginasSistema de Medición de Señales EEGCristian DallosAún no hay calificaciones
- 6.2 Técnicas de E.E.G (Electroencefalograma)Documento4 páginas6.2 Técnicas de E.E.G (Electroencefalograma)Dunia Nazareth Castillo GonzalezAún no hay calificaciones
- Bombas de InfusionDocumento30 páginasBombas de InfusionRoiser SanchezAún no hay calificaciones
- Breve Introducción A Las Técnicas de NeuroimagenDocumento26 páginasBreve Introducción A Las Técnicas de NeuroimagenBePe DenisseAún no hay calificaciones
- ELECTROCOCLEOGRAFIADocumento15 páginasELECTROCOCLEOGRAFIAGabriel CarrascoAún no hay calificaciones
- Potenciales Eferentes ValeDocumento3 páginasPotenciales Eferentes ValeVale TorresAún no hay calificaciones
- Campo Eléctrico. Detección de La Actividad Eléctrica Del Cerebro: ElectroencefalografíaDocumento8 páginasCampo Eléctrico. Detección de La Actividad Eléctrica Del Cerebro: ElectroencefalografíaConsuelo BahamondeAún no hay calificaciones
- Biofisica Apuntes 2°parcialDocumento16 páginasBiofisica Apuntes 2°parcialLorena!!Aún no hay calificaciones
- MielinizacionDocumento57 páginasMielinizacionVictoria Castillo100% (1)
- Actividad Electrica Del Sistema NerviosoDocumento16 páginasActividad Electrica Del Sistema NerviosoFernando Perez100% (2)
- Fisiologia de Transmision de Impulso NerviosoDocumento7 páginasFisiologia de Transmision de Impulso NerviosoIvantrosky MachukAún no hay calificaciones
- Técnicas de Registros PsicofisiológicosDocumento26 páginasTécnicas de Registros PsicofisiológicosMauricioIbáñezSánchezAún no hay calificaciones
- EQUIPOS DIAGNOSTICO - Electroencefalografo - Apuntes de Electromedicina Xavier PardellDocumento12 páginasEQUIPOS DIAGNOSTICO - Electroencefalografo - Apuntes de Electromedicina Xavier PardellRoberto Quinteiro GutierrezAún no hay calificaciones
- EmgDocumento16 páginasEmgMarioAún no hay calificaciones
- Biofisica 03Documento28 páginasBiofisica 03alexdestroyer250% (1)
- Comunicación IntraneuronalDocumento28 páginasComunicación IntraneuronalAnibal Urena RosarioAún no hay calificaciones
- Tarea 2 - Miguel Angel Ahumada Segura!!!Documento16 páginasTarea 2 - Miguel Angel Ahumada Segura!!!MIGUEL ANGEL AHUMADAAún no hay calificaciones
- BiopotencialesDocumento241 páginasBiopotencialesHéctor Alan AguilarAún no hay calificaciones
- Resumen TEMA 2 Técnicas Aplicadas en PsicobiologíaDocumento11 páginasResumen TEMA 2 Técnicas Aplicadas en PsicobiologíaNacho GutierrezAún no hay calificaciones
- Electro en Cefalo Graf oDocumento28 páginasElectro en Cefalo Graf oDany HernándezAún no hay calificaciones
- Exposición BioeléctricosDocumento68 páginasExposición BioeléctricosZenaida Núñez AlcántaraAún no hay calificaciones
- Fases Del SueñoDocumento9 páginasFases Del SueñoKaren HernándezAún no hay calificaciones
- Cap 10Documento38 páginasCap 10Francisco CermeñoAún no hay calificaciones
- Sistema Electrico Del Corazon y Ekg para BiomedicaDocumento50 páginasSistema Electrico Del Corazon y Ekg para BiomedicaDiana Carolina CameloAún no hay calificaciones
- EEG AlumnosDocumento106 páginasEEG AlumnosEloo Gm CiidAún no hay calificaciones
- 7) Electroencefalograma Humano-2015Documento7 páginas7) Electroencefalograma Humano-2015Sandra ZúñigaAún no hay calificaciones
- Tarea 2 - Trabajo ColaborativoDocumento46 páginasTarea 2 - Trabajo ColaborativoJuan Carlos Grisales MenesesAún no hay calificaciones
- Impulso Nervioso, Sinapsis y NeurotransmisoresDocumento17 páginasImpulso Nervioso, Sinapsis y NeurotransmisoresDagoberto Andres Fuentes Solis100% (2)
- Actividad Sistema Nervioso (1625)Documento12 páginasActividad Sistema Nervioso (1625)margarita gonzalezAún no hay calificaciones
- Guia Potencial de Accion y NeurotransmisoresDocumento11 páginasGuia Potencial de Accion y NeurotransmisoresAlvaro Andres Sanchez HurtadoAún no hay calificaciones
- Impulso Nervioso y SinapsisDocumento10 páginasImpulso Nervioso y SinapsisJesus García QuezadaAún no hay calificaciones
- 3 CLASE Tipos de Electrodos y DipoloDocumento31 páginas3 CLASE Tipos de Electrodos y DipoloPao spinaAún no hay calificaciones
- Tema 5 - Principales Técnicas Del Sistema Nervioso CentralDocumento7 páginasTema 5 - Principales Técnicas Del Sistema Nervioso CentralAnitta DFAún no hay calificaciones
- Sistema Nervioso Electroencefalograma (Eeg) PDFDocumento12 páginasSistema Nervioso Electroencefalograma (Eeg) PDFMilena PeraltaAún no hay calificaciones
- Tejido Nervioso T4-T4-T5Documento30 páginasTejido Nervioso T4-T4-T5Leon Diaz Ivan BaltazarAún no hay calificaciones
- Como Comunicarse Con El CerebroDocumento5 páginasComo Comunicarse Con El CerebroFlorencia MerinoAún no hay calificaciones
- EEFmapeoy NFCapitulorevisadofinalDocumento41 páginasEEFmapeoy NFCapitulorevisadofinalDamari Cardozo PizarroAún no hay calificaciones
- Partes de Un Casco de Estimulación TranscranealDocumento1 páginaPartes de Un Casco de Estimulación TranscranealHenry LagunaAún no hay calificaciones
- Practica de Laboratorio N0. 7Documento15 páginasPractica de Laboratorio N0. 7Edgar Osveli Temaj MacarioAún no hay calificaciones
- Tema 5 - ElectroneurografiaDocumento13 páginasTema 5 - ElectroneurografiaJunior FigueroaAún no hay calificaciones
- 3er Un BBCDocumento8 páginas3er Un BBCMarlene MorenoAún no hay calificaciones
- CA 6 Fisica Medica - En.espanolDocumento50 páginasCA 6 Fisica Medica - En.espanolCristhian Alexander Rafael BaylònAún no hay calificaciones
- 3 BioseñalesDocumento24 páginas3 BioseñalesAlexandro SuárezAún no hay calificaciones
- Los Métodos de Estudio Del Sistema NerviosoDocumento16 páginasLos Métodos de Estudio Del Sistema NerviosoÂngêl Rodrìguêz'Aún no hay calificaciones
- Las NeuronasDocumento5 páginasLas NeuronasLallypAún no hay calificaciones
- Señales BiopotencialesDocumento4 páginasSeñales BiopotencialesCAMILA MUNAR TORRESAún no hay calificaciones
- Medidas de Señales Bioelectricas, Clase de La Universidad Nacional Experimental FMDocumento43 páginasMedidas de Señales Bioelectricas, Clase de La Universidad Nacional Experimental FMLuis LopezAún no hay calificaciones
- Proyecto Dise oDocumento10 páginasProyecto Dise oFelipe MancillaAún no hay calificaciones
- TODOS Proponen 1 MínimoDocumento20 páginasTODOS Proponen 1 MínimoJob VillanuevaAún no hay calificaciones
- Electroencefalografía: Instituto Politécnico Nacional Escuela Nacional de Medicina Y HomeopatíaDocumento23 páginasElectroencefalografía: Instituto Politécnico Nacional Escuela Nacional de Medicina Y HomeopatíaMaría Rosa MartínezAún no hay calificaciones
- Reporte ProcesamientoDigitalSeñalesDocumento7 páginasReporte ProcesamientoDigitalSeñalesOmar MorenoAún no hay calificaciones
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaAún no hay calificaciones
- Fisiopatología ACVDocumento7 páginasFisiopatología ACVBrilli AnayaAún no hay calificaciones
- ESFENOIDESDocumento6 páginasESFENOIDESMartinSantaCruzAún no hay calificaciones
- CerebroDocumento17 páginasCerebrosophia perezAún no hay calificaciones
- 11 - e y F 1 - Torax y AbdomenDocumento55 páginas11 - e y F 1 - Torax y AbdomenJOSECARLOS RAMIREZ BREÑAAún no hay calificaciones
- Sistema Nervioso Central y Mascarilla FetalDocumento3 páginasSistema Nervioso Central y Mascarilla FetalYorky Saldaña TimanáAún no hay calificaciones
- Posiciones Anatómicas:: Posición AnatómicaDocumento16 páginasPosiciones Anatómicas:: Posición AnatómicaValentina MeierAún no hay calificaciones
- Anatomia Resumen CuelloDocumento8 páginasAnatomia Resumen Cuelloemanuel bonziAún no hay calificaciones
- HC VPLDocumento15 páginasHC VPLOber SánchezAún no hay calificaciones
- Evaluacion y Tratamiento Del Sistema NeuromiofascialDocumento2 páginasEvaluacion y Tratamiento Del Sistema NeuromiofascialDiana ExojoAún no hay calificaciones
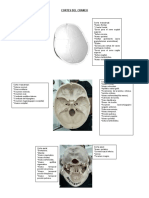
- Cortes Del CraneoDocumento3 páginasCortes Del CraneoIRAN ATZIRI ARTEAGA LIRAAún no hay calificaciones
- TiroidesDocumento18 páginasTiroidesapi-295455650Aún no hay calificaciones
- La Música en La Estimulación en El Cerebro de Los JóvenesDocumento7 páginasLa Música en La Estimulación en El Cerebro de Los JóvenesNelidaHuancaAún no hay calificaciones
- Pares Craneales - Es Slideshare Net 34Documento34 páginasPares Craneales - Es Slideshare Net 34Felipe Riestra0% (1)
- Presentacion Histologia Desarrollo de Cabeza y CuelloDocumento13 páginasPresentacion Histologia Desarrollo de Cabeza y CuelloAngelica NavtAún no hay calificaciones
- Brain y LenguajeDocumento12 páginasBrain y LenguajeZeptalanAún no hay calificaciones
- Eje CorticotropicoDocumento1 páginaEje CorticotropicoDalila DietschiAún no hay calificaciones
- Hormonas y EmbarazoDocumento61 páginasHormonas y EmbarazoTaly GarciaAún no hay calificaciones
- Me - 3.15 Area PeriocularDocumento28 páginasMe - 3.15 Area PeriocularLaura Miranda OjedaAún no hay calificaciones
- Los Ángulos de Movimiento de Las Articulaciones Del Miembro SuperiorDocumento2 páginasLos Ángulos de Movimiento de Las Articulaciones Del Miembro SuperiorMaría Fernanda SchmittAún no hay calificaciones
- Ficha AntropometricaDocumento7 páginasFicha Antropometricaviviana sanchezAún no hay calificaciones
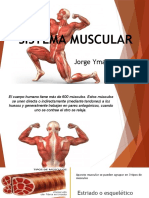
- Sistema-Muscular JyDocumento37 páginasSistema-Muscular JyDomelis Perez Vargas100% (2)
- Baremo LegalDocumento119 páginasBaremo LegalCecilia CardozoAún no hay calificaciones
- Plan de Entrenamiento FUERZADocumento2 páginasPlan de Entrenamiento FUERZAHector AngelesAún no hay calificaciones
- 5 Sentidos TripticoDocumento2 páginas5 Sentidos TripticoJulio Eustaquio Maguiña45% (11)
- PLEXOSDocumento19 páginasPLEXOSZaiira JiimenaAún no hay calificaciones
- Aorta Toracica - Abdominal - Iliacas Interna - ExternaDocumento55 páginasAorta Toracica - Abdominal - Iliacas Interna - ExternaLisbet CruzAún no hay calificaciones
- PlanimetriaDocumento12 páginasPlanimetriamaria Rambalde RamirezAún no hay calificaciones
- Crossfit y Entrenamiento FuncionalDocumento28 páginasCrossfit y Entrenamiento FuncionalGuitaryClimptonAún no hay calificaciones
- Fisiología Del CorazónDocumento2 páginasFisiología Del CorazónAndré GalloAún no hay calificaciones
- Anatomía Del MaxilarDocumento10 páginasAnatomía Del MaxilarMariana Rojas NuñezAún no hay calificaciones