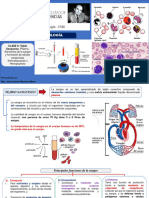
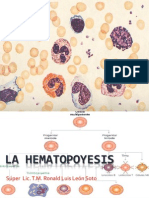
También podría gustarte
- Inmunología órganos y células del sistema inmuneDocumento9 páginasInmunología órganos y células del sistema inmuneHarold NavarreteAún no hay calificaciones
- T.I. Parásito U1Documento21 páginasT.I. Parásito U1Brayan PérezAún no hay calificaciones
- Células Granulocíticas Inmuno InnataDocumento3 páginasCélulas Granulocíticas Inmuno InnataTucurulan100% (1)
- Taller Histologia SangreDocumento10 páginasTaller Histologia Sangrekaren martinezAún no hay calificaciones
- Celulas y Organos Del Sistema InmuneDocumento4 páginasCelulas y Organos Del Sistema Inmuneliz reyesAún no hay calificaciones
- Resumen de Inmunologia - Celulas y Organos Del Sistema InmuneDocumento7 páginasResumen de Inmunologia - Celulas y Organos Del Sistema InmuneEsmeralda HernándezAún no hay calificaciones
- Marco Teórico HematoDocumento25 páginasMarco Teórico HematogramrealAún no hay calificaciones
- AGRANULOCITOSDocumento7 páginasAGRANULOCITOSnelson canquiAún no hay calificaciones
- Inmunología - Semana 2Documento18 páginasInmunología - Semana 2NahomiAún no hay calificaciones
- Células, Tejidos y Órganos Del Sistema InmuneDocumento12 páginasCélulas, Tejidos y Órganos Del Sistema InmuneRocío StürtzAún no hay calificaciones
- HematopoyesisDocumento19 páginasHematopoyesisJocelyne Calderon PadillaAún no hay calificaciones
- Inmunología 3Documento33 páginasInmunología 3CnlsAllAún no hay calificaciones
- Capturas de Las ClasesDocumento52 páginasCapturas de Las ClasesEdwin Nuñez DiazAún no hay calificaciones
- Medula OseaDocumento24 páginasMedula OseaMinerva García G.0% (1)
- Que Son Los Leucocitos InvestigacionDocumento20 páginasQue Son Los Leucocitos InvestigacionDocumentary SolutionsAún no hay calificaciones
- Sistema InmunologicoDocumento22 páginasSistema InmunologicoAnibal Antonio MenjivarAún no hay calificaciones
- Los Componentes Del Sistema InmunitarioDocumento11 páginasLos Componentes Del Sistema InmunitarioDaniela Garcés QuirogaAún no hay calificaciones
- Los Neutrófilos Normalmente Se Encuentran en El Torrente SanguíneoDocumento67 páginasLos Neutrófilos Normalmente Se Encuentran en El Torrente SanguíneoBianca Cristel Garduza CerdaAún no hay calificaciones
- G3s2imuno Células y Órganos Del Sistema InmuneDocumento6 páginasG3s2imuno Células y Órganos Del Sistema Inmunelucia mayancelaAún no hay calificaciones
- Organos y Tejidos Del Sistema InmuneDocumento6 páginasOrganos y Tejidos Del Sistema InmuneChristian GavilánezAún no hay calificaciones
- Células SanguíneasDocumento11 páginasCélulas SanguíneasCristian Uvidia SalgueroAún no hay calificaciones
- Arquitectura Del Sistema InmuneDocumento7 páginasArquitectura Del Sistema InmuneCarlos QuitianAún no hay calificaciones
- Paraclínicos BásicossDocumento8 páginasParaclínicos BásicossAndrea PaosulineAún no hay calificaciones
- CLASE 8 Tejido Sanguineo y HematopoyesisDocumento16 páginasCLASE 8 Tejido Sanguineo y HematopoyesisCésar Rodrigo Lope FloresAún no hay calificaciones
- Anatomía y Fisiología Del Sistema InmuneDocumento39 páginasAnatomía y Fisiología Del Sistema InmuneCésar Morante ChavarryAún no hay calificaciones
- TEMA 2. Células y órganos del sistema inmune y circulación linfocitariaDocumento22 páginasTEMA 2. Células y órganos del sistema inmune y circulación linfocitariaNina Souto BlancoAún no hay calificaciones
- Inmunidad Semana 1Documento11 páginasInmunidad Semana 1Guillermo Peralta LopezAún no hay calificaciones
- Organos LinfaticosDocumento18 páginasOrganos LinfaticosWraysthell CordovaAún no hay calificaciones
- Banco de Preguntas de Inmunología en Orden FINALDocumento12 páginasBanco de Preguntas de Inmunología en Orden FINALxlz100% (1)
- Fisiología Sistema InmuneDocumento60 páginasFisiología Sistema InmuneChristell AguilarAún no hay calificaciones
- Capitulo 7 MurrayDocumento4 páginasCapitulo 7 MurrayPedro A Hdz GalvanAún no hay calificaciones
- Sistema inmune humano: introducción a las células y órganosDocumento15 páginasSistema inmune humano: introducción a las células y órganosARLEIDYS RODRÍGUEZ RODRÍGUEZAún no hay calificaciones
- Clase 05 Hematopoyesis Celulas Maduras e InmadurasDocumento199 páginasClase 05 Hematopoyesis Celulas Maduras e InmadurasWilliam Apaza MamaniAún no hay calificaciones
- INMUNIDAD ADQUIRIDA O ESPECÍFICA TeoricoDocumento8 páginasINMUNIDAD ADQUIRIDA O ESPECÍFICA TeoricoPedro Insfran LiuzziAún no hay calificaciones
- LEUCOCITOSDocumento6 páginasLEUCOCITOSBrayan BarajasAún no hay calificaciones
- Sistema HematopoyeticoDocumento4 páginasSistema HematopoyeticoANTONIO PERDOMOAún no hay calificaciones
- Capítulo 2Documento12 páginasCapítulo 2Angie Pamela Mirella Colop PuacAún no hay calificaciones
- Tejidos LinfoidesDocumento33 páginasTejidos LinfoidesMishel Jordan ZeballosAún no hay calificaciones
- Sesión 1 CelulasDocumento5 páginasSesión 1 CelulasDaniel Duran MoenaAún no hay calificaciones
- Organos Linfaticos Primarios y SecundariosDocumento7 páginasOrganos Linfaticos Primarios y SecundariosItzel SalasAún no hay calificaciones
- HematopoyesisDocumento26 páginasHematopoyesisDalila TerniAún no hay calificaciones
- 04- MORFOFISIOLOGÍA DEL SISTEMA INMUNEDocumento6 páginas04- MORFOFISIOLOGÍA DEL SISTEMA INMUNECristAmuert 3Aún no hay calificaciones
- Tejido LinfoideDocumento6 páginasTejido LinfoideDiana RangelAún no hay calificaciones
- HEMATOPOYESIS Resumen KiaraDocumento6 páginasHEMATOPOYESIS Resumen KiarakiaraAún no hay calificaciones
- HEMATOPOYESISDocumento37 páginasHEMATOPOYESISEva Nelvi Guzman100% (1)
- Sangre AnemiasDocumento12 páginasSangre AnemiasELMER ELOY MAQUI SANTOAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Aula 2 - Celulas y Tejidos Del Sistema InmuneDocumento42 páginasAula 2 - Celulas y Tejidos Del Sistema InmuneLuana AraujoAún no hay calificaciones
- Tipos de Células InmunitariasDocumento3 páginasTipos de Células InmunitariasTomás Sánchez Roca100% (1)
- LINNFOPOYESISDocumento6 páginasLINNFOPOYESISChristian HidalgoAún no hay calificaciones
- Caracterìsticas y Funciones de Las Células Del SIDocumento107 páginasCaracterìsticas y Funciones de Las Células Del SISara Elizabeth CisnerosAún no hay calificaciones
- Células, Órganos Y Microambientes Del Sistema InmuneDocumento11 páginasCélulas, Órganos Y Microambientes Del Sistema InmuneJose Fernando Ortiz CarrilloAún no hay calificaciones
- Capitulo 7Documento18 páginasCapitulo 7SUGEILY NATALI MORALES LOPEZAún no hay calificaciones
- Organos Hematopoyeticos GrupalDocumento15 páginasOrganos Hematopoyeticos GrupalAnthony Fernando Arias MaldonadoAún no hay calificaciones
- Organos LinfoidesDocumento3 páginasOrganos LinfoidesEduardo FalcónAún no hay calificaciones
- Células sanguíneas: funciones y tiposDocumento13 páginasCélulas sanguíneas: funciones y tiposemontalvillo100% (3)
- Fisiología de La HematopoyesisDocumento4 páginasFisiología de La HematopoyesisAdhee VillaRiosAún no hay calificaciones
- Repaso del sistema inmune: células, órganos y respuestasDocumento73 páginasRepaso del sistema inmune: células, órganos y respuestasKarla Gonzalez DiazAún no hay calificaciones
- Histologia Medica LinfaticoDocumento4 páginasHistologia Medica LinfaticoAbraham Estrada sandovalAún no hay calificaciones
- Inmunologia Unidad 3Documento25 páginasInmunologia Unidad 3Cristhian DiazAún no hay calificaciones
- Claudia Ivette Rangel Martinez25062021Documento2 páginasClaudia Ivette Rangel Martinez25062021Claudia RangelAún no hay calificaciones
- Ensayo Hematologia-HematopoyesisDocumento14 páginasEnsayo Hematologia-HematopoyesisFran RetruzAún no hay calificaciones
- Felix Alejandro Quispe HernandezDocumento3 páginasFelix Alejandro Quispe HernandezJOEL ANTONY TORRES VILLENAAún no hay calificaciones
- Sistema InmunologicoDocumento19 páginasSistema InmunologicoTenorio Iparraguirre JulioAún no hay calificaciones
- PreguntasDocumento9 páginasPreguntasSerCL1Aún no hay calificaciones
- Cuestionario de NeutrofiloDocumento2 páginasCuestionario de NeutrofiloPatrick MartinezAún no hay calificaciones
- Valores Normales para Hemograma Por Grupo EtarioDocumento6 páginasValores Normales para Hemograma Por Grupo EtarioJrc65Aún no hay calificaciones
- Examen Resultado Unidades Valores de Referencia: QuímicaDocumento3 páginasExamen Resultado Unidades Valores de Referencia: QuímicaJARWIN CAICEDO CORTESAún no hay calificaciones
- Para PDF 2018 - Curso Formato - Chile - Recetas para Un Hemograma Bien HechoDocumento244 páginasPara PDF 2018 - Curso Formato - Chile - Recetas para Un Hemograma Bien HechoKarina Belén Espindola GuerraAún no hay calificaciones
- CAPÍTULO 2 - Células, Órganos y Microambientes Del Sistema InmuneDocumento55 páginasCAPÍTULO 2 - Células, Órganos y Microambientes Del Sistema InmuneDaniela MonroyAún no hay calificaciones
- Serie Roja EjerciciosDocumento6 páginasSerie Roja EjerciciosDiana V Landero CanteroAún no hay calificaciones
- Globulos BlancosDocumento5 páginasGlobulos BlancosBeralexa Transformadores PadmoundAún no hay calificaciones
- Fisiopatologia de Enfermedades AutoinmunesDocumento47 páginasFisiopatologia de Enfermedades AutoinmunesSociedad Científica de Estudiantes de Medicina de Ica93% (27)
- Interlucinas ?Documento15 páginasInterlucinas ?Exequiel GonzálezAún no hay calificaciones
- NERALIDADES1 Mecanismos InmunológicosDocumento29 páginasNERALIDADES1 Mecanismos InmunológicosWendy RodriguezAún no hay calificaciones
- Hematologia: Cuadro HematicoDocumento3 páginasHematologia: Cuadro Hematicojheison guevaraAún no hay calificaciones
- Linfocitos TDocumento22 páginasLinfocitos TSamAún no hay calificaciones
- Cuestionario 1 FisioDocumento3 páginasCuestionario 1 FisioLu RAún no hay calificaciones
- HEMATOPOYESISDocumento78 páginasHEMATOPOYESISCindy Flores TipteAún no hay calificaciones
- Cuadro Comparativo de Células de La Sangre - Alanís LozanoDocumento2 páginasCuadro Comparativo de Células de La Sangre - Alanís Lozano样样100% (1)
- Histologia I - SANGREDocumento30 páginasHistologia I - SANGREdouglasrodrydasilvaAún no hay calificaciones
- Práctica 3 - Tejido SanguíneoDocumento61 páginasPráctica 3 - Tejido SanguíneoRichard CabelloAún no hay calificaciones
- Resultados de análisis clínicos de Jacobo López GarcíaDocumento1 páginaResultados de análisis clínicos de Jacobo López Garcíaalejandro ucAún no hay calificaciones
- Páginas Extraídas de Ross Histologia Texto y Atlas 7a EdDocumento4 páginasPáginas Extraídas de Ross Histologia Texto y Atlas 7a EdCarlosAún no hay calificaciones
- Max HemogramaDocumento2 páginasMax HemogramaEnrique Hernández SolisAún no hay calificaciones
- Maduración (Ontogenia) de Linfocitos T - Selección Positiva y Negativa - Compromiso de LíneaDocumento12 páginasMaduración (Ontogenia) de Linfocitos T - Selección Positiva y Negativa - Compromiso de LíneaJhonny M CubaAún no hay calificaciones
- Recuento Mínimo de Leucocitos para Realizar La Fórmula Leucocitaria en Frotis de Sangre PeriféricaDocumento20 páginasRecuento Mínimo de Leucocitos para Realizar La Fórmula Leucocitaria en Frotis de Sangre PeriféricaLuis Carlos Quinto CuzcanoAún no hay calificaciones
- F. Salud 9-3 - Tercera UnidadDocumento49 páginasF. Salud 9-3 - Tercera UnidadChristian CasusolAún no hay calificaciones
- Hematopoyesis 1Documento35 páginasHematopoyesis 1Ruben Rojas RAún no hay calificaciones
- Resultados de LaboratorioDocumento5 páginasResultados de LaboratorioMIGUEL ANGELAún no hay calificaciones