También podría gustarte
- Curso Cosmetica Solida Natural - 1Documento38 páginasCurso Cosmetica Solida Natural - 1FRANCISCA REYES MARTINEZ100% (11)
- Taller Procesos Bioelectricos Ulibre 2020Documento6 páginasTaller Procesos Bioelectricos Ulibre 2020Valentina Moreno VivasAún no hay calificaciones
- Ensayo de Potencial de AcciónDocumento3 páginasEnsayo de Potencial de AcciónAndrea HernandezAún no hay calificaciones
- Síntesis Del OctenoDocumento4 páginasSíntesis Del OctenoAndres Felipe LealAún no hay calificaciones
- Experimento de Laboratorio Con NERVEDocumento16 páginasExperimento de Laboratorio Con NERVENicoll Perez ManosalvaAún no hay calificaciones
- Mega ResumenFISIOLOGIADocumento57 páginasMega ResumenFISIOLOGIAkirkoxisAún no hay calificaciones
- Netter Fundamentos de Fisiologia - LEONES - POR - LA - SALUDf 5Documento1 páginaNetter Fundamentos de Fisiologia - LEONES - POR - LA - SALUDf 5shine lighAún no hay calificaciones
- Potencial de MembranaDocumento6 páginasPotencial de MembranagustafAún no hay calificaciones
- Parcial 3 (Bioelectricidad)Documento16 páginasParcial 3 (Bioelectricidad)Nicole Carolina Quintero RomeroAún no hay calificaciones
- TP 4 PropiedadespasivasdelamembranaDocumento10 páginasTP 4 PropiedadespasivasdelamembranaVero OrtegaAún no hay calificaciones
- Fisiologia Neuromuscular 2012Documento63 páginasFisiologia Neuromuscular 2012Luis Alberto Isea MAún no hay calificaciones
- Experimento de Laboratorio Con NERVEDocumento18 páginasExperimento de Laboratorio Con NERVENicoll Perez ManosalvaAún no hay calificaciones
- Rivolta Miguel y Lucas Benavides 2016.apunte de Cátedra Unidad 5. Bases Físicas de Los Fenómenos BioeléctricosDocumento12 páginasRivolta Miguel y Lucas Benavides 2016.apunte de Cátedra Unidad 5. Bases Físicas de Los Fenómenos BioeléctricosJennifer MarcatelliAún no hay calificaciones
- Vídeos Explicación Unidad Iii (Fisiología)Documento6 páginasVídeos Explicación Unidad Iii (Fisiología)Amy Lilian Verdooren UzcateguiAún no hay calificaciones
- Clases - Fisiología CardiovascularDocumento76 páginasClases - Fisiología CardiovascularEstefania Gonzalez100% (1)
- Seminario Biopotenciales FisiologiaDocumento9 páginasSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- Simulacion Por ComputadoraDocumento17 páginasSimulacion Por ComputadoraJeeniffer CookieeAún no hay calificaciones
- Potenciales Bioeléctricos USEP 2022Documento28 páginasPotenciales Bioeléctricos USEP 2022YESENIA SARAIT AGUILAR HERRERAAún no hay calificaciones
- Fisiologia, PotencialesDocumento17 páginasFisiologia, PotencialesAnthony Namay PoloAún no hay calificaciones
- Impulso Nervioso (Potencial de Acción)Documento6 páginasImpulso Nervioso (Potencial de Acción)Cesar DuqueAún no hay calificaciones
- 1-Principios Basicos de Electrofisiologia CardiacaDocumento9 páginas1-Principios Basicos de Electrofisiologia CardiacaJustina MuttiAún no hay calificaciones
- Resumen Examen NeurofisioDocumento62 páginasResumen Examen NeurofisioJuan Pablo Díaz BecerraAún no hay calificaciones
- TP Potenciales y Sinapsis PDFDocumento10 páginasTP Potenciales y Sinapsis PDFSabrina PortelliAún no hay calificaciones
- 01 ExcitabilidadDocumento48 páginas01 ExcitabilidadCosmito FulanitoAún no hay calificaciones
- BiopotencialesDocumento6 páginasBiopotencialesLiseth100% (1)
- BIOFISICADocumento55 páginasBIOFISICAJanet Ordinola100% (1)
- Electrofisiologia GeneralDocumento5 páginasElectrofisiologia GeneralFlavinho YrigoyenAún no hay calificaciones
- Biofisica de Las Membranas ExcitablesDocumento19 páginasBiofisica de Las Membranas ExcitablesYamil Valencia100% (1)
- Circuito EquivalenteDocumento12 páginasCircuito EquivalenteKaren GonzalezAún no hay calificaciones
- 2 - Tejidos ExcitablesDocumento11 páginas2 - Tejidos ExcitablesMartin TaraffoAún no hay calificaciones
- Gradientes Iónicos y Actividad BioeléctricaDocumento4 páginasGradientes Iónicos y Actividad BioeléctricaRaúl Zapater LópezAún no hay calificaciones
- Tema 2Documento36 páginasTema 2Citni22Aún no hay calificaciones
- Tema 3. Equilibrio Ionico y Potencial de Membrana de ReposoDocumento26 páginasTema 3. Equilibrio Ionico y Potencial de Membrana de ReposoXimenna Antuaneth TapiaAún no hay calificaciones
- Conduccion Del Impulso NerviosoDocumento17 páginasConduccion Del Impulso NerviosoomarcvetAún no hay calificaciones
- Clase n5 NbaDocumento7 páginasClase n5 NbaFlorencia9 GhioAún no hay calificaciones
- Capitulo 3 - Neurociencia PDFDocumento16 páginasCapitulo 3 - Neurociencia PDFDavid RojasAún no hay calificaciones
- Electrofisiología Del EKGDocumento67 páginasElectrofisiología Del EKGHans M. Valdiviezo BonifazAún no hay calificaciones
- FisiologíaDocumento4 páginasFisiologíaMartha PadillaAún no hay calificaciones
- Control de Los Patrones de Descarga en El SomaDocumento3 páginasControl de Los Patrones de Descarga en El SomaSTEVEN WILTER JUNIOR MENDEZ ALVARADOAún no hay calificaciones
- Guía de Seminario 3 Excitabilidad, Potencial de Acción y Sinapsis.Documento13 páginasGuía de Seminario 3 Excitabilidad, Potencial de Acción y Sinapsis.zScratAún no hay calificaciones
- Bioelectric I DadDocumento5 páginasBioelectric I DadAriana Figueroa OviedoAún no hay calificaciones
- Metodo de Prediccion - TesisRicardalast-fDocumento30 páginasMetodo de Prediccion - TesisRicardalast-fAxel David Castro AguilarAún no hay calificaciones
- Fisiología R1 CardiologíaDocumento9 páginasFisiología R1 CardiologíaGabrielly RobledilhoAún no hay calificaciones
- Teorico ElectorfisiologiaDocumento9 páginasTeorico ElectorfisiologiaIaraAún no hay calificaciones
- Fenomenos BioelectricosDocumento4 páginasFenomenos Bioelectricosnicolas villegasAún no hay calificaciones
- Resumen Kandel 1Documento122 páginasResumen Kandel 1DavidAún no hay calificaciones
- BIOPOTENCIALES FisiologíaDocumento6 páginasBIOPOTENCIALES FisiologíaDJ EDWAún no hay calificaciones
- M3 Qué DecirDocumento7 páginasM3 Qué DecirRaïssaDomonAún no hay calificaciones
- Bioelectricidad UsmpDocumento42 páginasBioelectricidad UsmpedgarAún no hay calificaciones
- Resumen PurvesDocumento64 páginasResumen PurvesBrigitte MaldonadoAún no hay calificaciones
- Ley de KirchoffDocumento5 páginasLey de KirchoffFrancisco Javier Ramirez ChavezAún no hay calificaciones
- Factores Que Intervienen en El Desarrollo Del Potencial de AcciónDocumento6 páginasFactores Que Intervienen en El Desarrollo Del Potencial de AcciónCecilia PachecoAún no hay calificaciones
- 10 - La PsicobiologíaDocumento5 páginas10 - La PsicobiologíaWeb PublicidadAún no hay calificaciones
- PROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClaseDocumento10 páginasPROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClasewpmariaantoniaAún no hay calificaciones
- Propiedades Eléctricas de Las Membranas de Las Células ExcitablesDocumento5 páginasPropiedades Eléctricas de Las Membranas de Las Células ExcitablesCésar Yael Partida Pérez50% (2)
- TipeoDocumento32 páginasTipeoMartin QuezadaAún no hay calificaciones
- TEMA 5-Potencial de AcciónDocumento11 páginasTEMA 5-Potencial de AcciónAlba Martinez MorenoAún no hay calificaciones
- Cardio TotalDocumento125 páginasCardio TotalBrenda PusdáAún no hay calificaciones
- Experiencias sobre circuitos eléctricos en serie operados por corriente directaDe EverandExperiencias sobre circuitos eléctricos en serie operados por corriente directaAún no hay calificaciones
- Teoría electromagnética para estudiantes de ingeniería: Notas de claseDe EverandTeoría electromagnética para estudiantes de ingeniería: Notas de claseCalificación: 4.5 de 5 estrellas4.5/5 (7)
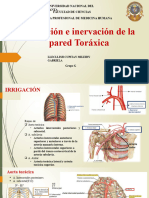
- Seminario 1 Irrigación e Inervación Del ToráxDocumento8 páginasSeminario 1 Irrigación e Inervación Del ToráxGabriela JaramilloAún no hay calificaciones
- Cronograma Producto Y, RR-bioquimica 2024Documento12 páginasCronograma Producto Y, RR-bioquimica 2024Gabriela JaramilloAún no hay calificaciones
- Silabo Bioquímica UNS 2024-1 Final FDocumento12 páginasSilabo Bioquímica UNS 2024-1 Final FGabriela JaramilloAún no hay calificaciones
- Configuración InternaDocumento17 páginasConfiguración InternaGabriela JaramilloAún no hay calificaciones
- Pericardio y CorazonDocumento26 páginasPericardio y CorazonGabriela JaramilloAún no hay calificaciones
- Examen Mental Adulto MayorDocumento14 páginasExamen Mental Adulto MayorGabriela JaramilloAún no hay calificaciones
- Comunicación - LA MONOGRAFIADocumento3 páginasComunicación - LA MONOGRAFIAGabriela JaramilloAún no hay calificaciones
- Universidad Dr. José Matías Delgado Red Bibliotecaria Matías Derechos de PublicaciónDocumento82 páginasUniversidad Dr. José Matías Delgado Red Bibliotecaria Matías Derechos de PublicaciónKugoleoAún no hay calificaciones
- Sikalastic-1c HOJA SEGDocumento9 páginasSikalastic-1c HOJA SEGBessyAún no hay calificaciones
- Degradación de Ácido Poliláctico Hidroxiapatita y Ácido Poliglicólico en Fluido Corporal SimuladoDocumento7 páginasDegradación de Ácido Poliláctico Hidroxiapatita y Ácido Poliglicólico en Fluido Corporal SimuladoKarina CabezasAún no hay calificaciones
- Molienda Caracterización y Equipos de OperaciónDocumento10 páginasMolienda Caracterización y Equipos de OperaciónDANIEL FRANCISCO BOCANEGRA ROJASAún no hay calificaciones
- Universidad de CartagenaDocumento7 páginasUniversidad de CartagenaCristian Ronco PerezAún no hay calificaciones
- República Bolivariana de VenezuelaDocumento17 páginasRepública Bolivariana de VenezuelaIrmary EspinozaAún no hay calificaciones
- Catálogo Kosmoflex 2023 Baja ResoluciónDocumento40 páginasCatálogo Kosmoflex 2023 Baja Resoluciónpatricio122587Aún no hay calificaciones
- Plantas-Medicinales VITAMINASDocumento5 páginasPlantas-Medicinales VITAMINASkaren turpoAún no hay calificaciones
- Conclusiones - Investigación Caso "Peñoles"Documento15 páginasConclusiones - Investigación Caso "Peñoles"RA AlcocerAún no hay calificaciones
- Antagonistas Colinergicos 1Documento4 páginasAntagonistas Colinergicos 1Andy GarciaAún no hay calificaciones
- Practica N 6 ResueltoDocumento6 páginasPractica N 6 ResueltoJennifer ChávezAún no hay calificaciones
- Biomembranas y Transporte CelularDocumento7 páginasBiomembranas y Transporte CelularADRIAN SANCHEZ CASTILLOAún no hay calificaciones
- 14 Esterilizacion Del Material EmpleadoDocumento5 páginas14 Esterilizacion Del Material Empleadomilirosi25111Aún no hay calificaciones
- Jee93 0180Documento9 páginasJee93 0180pasanteglobalchemAún no hay calificaciones
- Refractarios de CarbónDocumento7 páginasRefractarios de CarbónRafaelAún no hay calificaciones
- Removedor de PinturaDocumento16 páginasRemovedor de PinturaElkin MesaAún no hay calificaciones
- Sellador Acrilico AnypsaDocumento2 páginasSellador Acrilico AnypsaKevin Paul Arca RojasAún no hay calificaciones
- Alquenos y AlquinosDocumento20 páginasAlquenos y Alquinos0Aún no hay calificaciones
- Membranas en PVC GDocumento4 páginasMembranas en PVC GDanyAún no hay calificaciones
- Semana 3 - Informe 3 - Componentes No Protoplamaticos SecundariosDocumento8 páginasSemana 3 - Informe 3 - Componentes No Protoplamaticos SecundariosSebastián García TerronesAún no hay calificaciones
- Mod03-Lec01 - Cromatografia de Intercambio IonicoDocumento4 páginasMod03-Lec01 - Cromatografia de Intercambio IonicoMario Antonio Araya MorosoAún no hay calificaciones
- Hds Plastificante 325Documento6 páginasHds Plastificante 325Gestión AmbientalAún no hay calificaciones
- Informe AditivosDocumento10 páginasInforme AditivosJefferson LlangariAún no hay calificaciones
- Pérez Cruz David - Determinación Ácido Tartárico en VinoDocumento7 páginasPérez Cruz David - Determinación Ácido Tartárico en VinoDAVID EMMANUEL PEREZ CRUZAún no hay calificaciones
- Obtencion y Caracterizacion de Quitina y Quitosano Del Emerita Analoga A Escala PilotoDocumento16 páginasObtencion y Caracterizacion de Quitina y Quitosano Del Emerita Analoga A Escala Pilotojorge luis maldonadoAún no hay calificaciones
- Oxido de Zinc Hoja de SeguridadDocumento5 páginasOxido de Zinc Hoja de SeguridadDuvan GonzalezAún no hay calificaciones
- Problemario de Balance2000Documento32 páginasProblemario de Balance2000Montserrat VigilAún no hay calificaciones
- Soluciones (9 12)Documento5 páginasSoluciones (9 12)Gabriel Vigo JaraAún no hay calificaciones