También podría gustarte
- Fenomenos BioelectricosDocumento4 páginasFenomenos Bioelectricosnicolas villegasAún no hay calificaciones
- Potencial de Accion y MembranaDocumento8 páginasPotencial de Accion y MembranaFrancisco MoroyoquiAún no hay calificaciones
- BIOFISICADocumento55 páginasBIOFISICAJanet Ordinola100% (1)
- Tema 6 Pasado A LimpioDocumento6 páginasTema 6 Pasado A LimpioNerea Rodríguez GómezAún no hay calificaciones
- Tema 3. Potencial de MembranaDocumento11 páginasTema 3. Potencial de MembranaAlbaAún no hay calificaciones
- Resumen PurvesDocumento64 páginasResumen PurvesBrigitte MaldonadoAún no hay calificaciones
- La Generación y La Transmisión Del Impulso NerviosoDocumento21 páginasLa Generación y La Transmisión Del Impulso NerviosomixoneqAún no hay calificaciones
- BIOPOTENCIALES FisiologíaDocumento6 páginasBIOPOTENCIALES FisiologíaDJ EDWAún no hay calificaciones
- 10 - La PsicobiologíaDocumento5 páginas10 - La PsicobiologíaWeb PublicidadAún no hay calificaciones
- Clase n5 NbaDocumento7 páginasClase n5 NbaFlorencia9 GhioAún no hay calificaciones
- Tema 2Documento36 páginasTema 2Citni22Aún no hay calificaciones
- La Comunicación NeuronalDocumento11 páginasLa Comunicación NeuronalMarianne DaswoodAún no hay calificaciones
- Potenciales de Memrana y Potenciales de AcciónDocumento17 páginasPotenciales de Memrana y Potenciales de AcciónGabriela Hu MaAún no hay calificaciones
- Excitacion CelularDocumento28 páginasExcitacion CelularGerson Loyola Javes100% (1)
- Potencial de MembranaDocumento6 páginasPotencial de MembranagustafAún no hay calificaciones
- Tema 3. Equilibrio Ionico y Potencial de Membrana de ReposoDocumento26 páginasTema 3. Equilibrio Ionico y Potencial de Membrana de ReposoXimenna Antuaneth TapiaAún no hay calificaciones
- Impulso Nervioso (Potencial de Acción)Documento6 páginasImpulso Nervioso (Potencial de Acción)Cesar DuqueAún no hay calificaciones
- Ensayo de Potencial de AcciónDocumento3 páginasEnsayo de Potencial de AcciónAndrea HernandezAún no hay calificaciones
- Potencial de MembranaDocumento3 páginasPotencial de MembranaJoel DuarteAún no hay calificaciones
- Seminario Biopotenciales FisiologiaDocumento9 páginasSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- ApuntesNeurofisio ResaltadoDocumento128 páginasApuntesNeurofisio ResaltadoFabian CesarAún no hay calificaciones
- Canales Iónicos y Potenciales MembranaDocumento17 páginasCanales Iónicos y Potenciales MembranaAnthony Namay PoloAún no hay calificaciones
- Potencial de DifusiónDocumento4 páginasPotencial de DifusiónAriana Reliche Briones50% (2)
- 3- Potenciales de Membrana y Potenciales de AcciónDocumento63 páginas3- Potenciales de Membrana y Potenciales de Accióndr.josefernandezlAún no hay calificaciones
- TEJIDOS EXCITABLES Guia de Estudio PDFDocumento42 páginasTEJIDOS EXCITABLES Guia de Estudio PDFMaria Luz VirasoroAún no hay calificaciones
- Seminario de PotencialesDocumento10 páginasSeminario de PotencialesKatherine CAAún no hay calificaciones
- Excitabilidad CelularDocumento30 páginasExcitabilidad CelularFernando PazAún no hay calificaciones
- Vídeos Explicación Unidad Iii (Fisiología)Documento6 páginasVídeos Explicación Unidad Iii (Fisiología)Amy Lilian Verdooren UzcateguiAún no hay calificaciones
- Potenciales membrana y transporte iónicoDocumento62 páginasPotenciales membrana y transporte iónicoJuan Pablo Díaz BecerraAún no hay calificaciones
- Bases Biofisicas de Fenomenos EléctricosDocumento14 páginasBases Biofisicas de Fenomenos EléctricosDiego Sanchez PerezAún no hay calificaciones
- Cardio TotalDocumento125 páginasCardio TotalBrenda PusdáAún no hay calificaciones
- B 2 BacaDocumento32 páginasB 2 Bacajgfb180165Aún no hay calificaciones
- Excitabilidad Celular - MEHU 632Documento24 páginasExcitabilidad Celular - MEHU 632ARIANA GREEIS LUJAN REBAZAAún no hay calificaciones
- BiopotencialesDocumento6 páginasBiopotencialesLiseth100% (1)
- Biofísica y NeuronasDocumento11 páginasBiofísica y Neuronasanonymo0% (1)
- Conducta Eléctrica de La MembranaDocumento68 páginasConducta Eléctrica de La Membranajuarddan100% (1)
- Origen Del Potencial de Membrana en Reposo NormalDocumento3 páginasOrigen Del Potencial de Membrana en Reposo NormalBrithey SanchezAún no hay calificaciones
- Potenciales de MembranaDocumento36 páginasPotenciales de Membranapepe_074Aún no hay calificaciones
- Clase n5 NbaDocumento8 páginasClase n5 NbaFlorencia9 GhioAún no hay calificaciones
- Fisiología neuronalDocumento30 páginasFisiología neuronalGadiel Couoh Rosado100% (1)
- PROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClaseDocumento10 páginasPROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClasewpmariaantoniaAún no hay calificaciones
- Netter Fundamentos de Fisiologia - LEONES - POR - LA - SALUDf 5Documento1 páginaNetter Fundamentos de Fisiologia - LEONES - POR - LA - SALUDf 5shine lighAún no hay calificaciones
- 1-Principios Basicos de Electrofisiologia CardiacaDocumento9 páginas1-Principios Basicos de Electrofisiologia CardiacaJustina MuttiAún no hay calificaciones
- RepasoDocumento14 páginasRepasoMiriam Ávila ValladaresAún no hay calificaciones
- Bases de la comunicación neuronalDocumento35 páginasBases de la comunicación neuronalGaia OrevAún no hay calificaciones
- Potencial membrana reposoDocumento11 páginasPotencial membrana reposoMiriam Ávila ValladaresAún no hay calificaciones
- Potencial de AccionDocumento5 páginasPotencial de Accionjairo santiagoAún no hay calificaciones
- Origen biopotencialesDocumento28 páginasOrigen biopotencialespedroAún no hay calificaciones
- Tema 3 DesarrolloDocumento16 páginasTema 3 Desarrolloantonio garcia martinezAún no hay calificaciones
- TipeoDocumento32 páginasTipeoMartin QuezadaAún no hay calificaciones
- Actividad Eléctrica en Los AxonesDocumento33 páginasActividad Eléctrica en Los AxonesGustavo Peña100% (1)
- 10Documento25 páginas10D BastianAún no hay calificaciones
- Potencial de Membrana y Potencial D Accion-2Documento48 páginasPotencial de Membrana y Potencial D Accion-2wilson clavijoAún no hay calificaciones
- Corazon Como BombaDocumento65 páginasCorazon Como BombaJCAún no hay calificaciones
- Potenciales de Membrana y Potenciales de AcciónDocumento31 páginasPotenciales de Membrana y Potenciales de AcciónAnto FloresAún no hay calificaciones
- Psicología Biológica. Rosenzweing 3Documento3 páginasPsicología Biológica. Rosenzweing 3Agustín PeruginiAún no hay calificaciones
- 2 (1) (1) ClasePotencialdeMembranaDocumento21 páginas2 (1) (1) ClasePotencialdeMembranaBastian Rodrigo Almendra FernandezAún no hay calificaciones
- Teoría electromagnética para estudiantes de ingeniería: Notas de claseDe EverandTeoría electromagnética para estudiantes de ingeniería: Notas de claseCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Curso de enfermería básica: Clase 2 sobre dosis, vías de administración y traumatismosDocumento22 páginasCurso de enfermería básica: Clase 2 sobre dosis, vías de administración y traumatismosMartin TaraffoAún no hay calificaciones
- Chopita - 7 Genetica MendelianaDocumento7 páginasChopita - 7 Genetica MendelianaMartin TaraffoAún no hay calificaciones
- Curso de Enfermeria 2016 Clase 3Documento30 páginasCurso de Enfermeria 2016 Clase 3Martin TaraffoAún no hay calificaciones
- Curso de Enfermeria 2016 Clase 1Documento23 páginasCurso de Enfermeria 2016 Clase 1Martin TaraffoAún no hay calificaciones
- Chopita - 4 Biotecnologia y Genomica AnimalDocumento7 páginasChopita - 4 Biotecnologia y Genomica AnimalMartin TaraffoAún no hay calificaciones
- Chopita - 0-TECNICAS-DIAGNOSTICASDocumento15 páginasChopita - 0-TECNICAS-DIAGNOSTICASMartin TaraffoAún no hay calificaciones
- Chopita - 5-ABORTIGENASDocumento12 páginasChopita - 5-ABORTIGENASMartin TaraffoAún no hay calificaciones
- Enfermedades-crónicas-tuberculosisDocumento12 páginasEnfermedades-crónicas-tuberculosisMartin TaraffoAún no hay calificaciones
- Chopita - Cuadro BrucelosisDocumento4 páginasChopita - Cuadro BrucelosisMartin TaraffoAún no hay calificaciones
- Chopita - 3-ENTERITOGENICAS Parte 2Documento7 páginasChopita - 3-ENTERITOGENICAS Parte 2Martin TaraffoAún no hay calificaciones
- Chopita - 2-PIOGENASDocumento14 páginasChopita - 2-PIOGENASMartin TaraffoAún no hay calificaciones
- Chopita - 2 CromatinaDocumento11 páginasChopita - 2 CromatinaMartin TaraffoAún no hay calificaciones
- Diferenciales de enfermedades abortígenas en especies productorasDocumento2 páginasDiferenciales de enfermedades abortígenas en especies productorasMartin TaraffoAún no hay calificaciones
- Chopita - 1-TOXOINFECCIOSASDocumento15 páginasChopita - 1-TOXOINFECCIOSASMartin TaraffoAún no hay calificaciones
- Introducción a las enfermedades infecciosasDocumento14 páginasIntroducción a las enfermedades infecciosasFernanda GaiadAún no hay calificaciones
- Chopita - 6 Ciclo CelularDocumento6 páginasChopita - 6 Ciclo CelularMartin TaraffoAún no hay calificaciones
- Chopita - Cardio 1Documento5 páginasChopita - Cardio 1Martin TaraffoAún no hay calificaciones
- Chopita - 3 Regulacion de La Expresion GenicaDocumento6 páginasChopita - 3 Regulacion de La Expresion GenicaMartin TaraffoAún no hay calificaciones
- Wong Kiew Kit - El Gran Libro de La Medicina ChinaDocumento332 páginasWong Kiew Kit - El Gran Libro de La Medicina Chinaliilu96% (26)
- Chopita - Cardio 3Documento9 páginasChopita - Cardio 3Martin TaraffoAún no hay calificaciones
- Chopita - 5 Marcadores MolecularesDocumento4 páginasChopita - 5 Marcadores MolecularesMartin TaraffoAún no hay calificaciones
- Chopita - 1 Genes y GenomasDocumento4 páginasChopita - 1 Genes y GenomasMartin TaraffoAún no hay calificaciones
- Chopita - Acido BaseDocumento6 páginasChopita - Acido BaseMartin TaraffoAún no hay calificaciones
- 3 - SNA ReceptoresDocumento2 páginas3 - SNA ReceptoresMartin TaraffoAún no hay calificaciones
- Clinica VeterinariaDocumento35 páginasClinica VeterinariaMartin Taraffo100% (3)
- Situacion y Perspectiva Del Sector AgropecuarioDocumento13 páginasSituacion y Perspectiva Del Sector AgropecuarioMartin TaraffoAún no hay calificaciones
- Organizacion EmpresaDocumento38 páginasOrganizacion EmpresaMartin TaraffoAún no hay calificaciones
- Clinica VeterinariaDocumento35 páginasClinica VeterinariaMartin Taraffo100% (3)
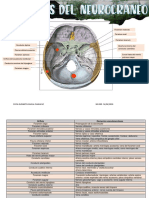
- Orificios Del NeurocraneoDocumento2 páginasOrificios Del NeurocraneoSofia MaflaAún no hay calificaciones
- Sistema nervioso y sus funcionesDocumento4 páginasSistema nervioso y sus funcionesGerard Sorto100% (1)
- Equine Internal Medicine MUSCULOESQUELETICO - En.esDocumento34 páginasEquine Internal Medicine MUSCULOESQUELETICO - En.esDamian Chicaiza100% (1)
- 3ro. Año de Educación Secundaria Comunitaria ProductivaDocumento10 páginas3ro. Año de Educación Secundaria Comunitaria ProductivaAshly Meredith Gomez GuardiaAún no hay calificaciones
- Resumen U 1,2,3,4,5 Neuro 1Documento31 páginasResumen U 1,2,3,4,5 Neuro 1IaraCamila GuerraAún no hay calificaciones
- Historia de La NeurocienciaDocumento6 páginasHistoria de La NeurocienciaNicole BastidasAún no hay calificaciones
- Clase 9 Lunes 24 (2) Estructura IDocumento79 páginasClase 9 Lunes 24 (2) Estructura IKiara Jimena Vidalon VenturaAún no hay calificaciones
- Bases Neurologicas Del Mov Humano UnmsmDocumento113 páginasBases Neurologicas Del Mov Humano UnmsmJ Mikel NinhoAún no hay calificaciones
- Psicologia AnaDocumento10 páginasPsicologia AnaCARLOS ARTURO GUTIERREZ GOMEZAún no hay calificaciones
- 01111111111111Documento25 páginas01111111111111Bladimir F J MendezAún no hay calificaciones
- Resolucion A La Guia de Estudio Unidad I Sistema NerviosoDocumento12 páginasResolucion A La Guia de Estudio Unidad I Sistema Nerviosoyesica yamila escuderoAún no hay calificaciones
- Sistema NerviosoDocumento21 páginasSistema Nerviosocelenne villanuevaAún no hay calificaciones
- Aspectos Historicos de La NeurologiaDocumento5 páginasAspectos Historicos de La NeurologiaYorisbel De La CruzAún no hay calificaciones
- Terminología Medica - Sistema NerviosoDocumento26 páginasTerminología Medica - Sistema NerviosoJose Benjamin Almanza Orozco100% (1)
- La Funcion Nerviosa OctavosDocumento5 páginasLa Funcion Nerviosa OctavosJaime Hernando Ayala ChavezAún no hay calificaciones
- Sistema Nervioso PisanoDocumento34 páginasSistema Nervioso PisanonicolasAún no hay calificaciones
- 3 InjertosDocumento21 páginas3 InjertosClaudio R. MonteroAún no hay calificaciones
- Sistema Nervioso y Org. de Los SentidosDocumento63 páginasSistema Nervioso y Org. de Los SentidosJasna ZovakAún no hay calificaciones
- Terapia Neural HunekeDocumento29 páginasTerapia Neural HunekejlgallardoAún no hay calificaciones
- Apunte-Inervación Del Sistema GastrointestinalDocumento9 páginasApunte-Inervación Del Sistema GastrointestinalBren MtzzAún no hay calificaciones
- Tarea 1 Sistema Nervioso y SentidosDocumento4 páginasTarea 1 Sistema Nervioso y SentidosSugey MolinaAún no hay calificaciones
- Preguntas NeuroDocumento17 páginasPreguntas NeuroClaudia GamboaAún no hay calificaciones
- Anestésicos LocalesDocumento8 páginasAnestésicos Localesjoselinne009Aún no hay calificaciones
- Osteopatía estructural guía de estudio FEMATEC TáchiraDocumento57 páginasOsteopatía estructural guía de estudio FEMATEC TáchiraLandy PerezAún no hay calificaciones
- Clasificación Del Sistema NerviosoDocumento14 páginasClasificación Del Sistema NerviosoLeydi VargasAún no hay calificaciones
- POLINEUROPATÍASDocumento49 páginasPOLINEUROPATÍASIvan UrizaAún no hay calificaciones
- Guía de Aprendizaje Sistema Nervioso Grado 9Documento7 páginasGuía de Aprendizaje Sistema Nervioso Grado 9Victor MazoAún no hay calificaciones
- Taller de Valoracion Sobre El Sistema NerviosoDocumento13 páginasTaller de Valoracion Sobre El Sistema NerviosoDeicy Carolina Galvis PeñaAún no hay calificaciones
- Diapositivas Nervioso Central y Periferico 1Documento28 páginasDiapositivas Nervioso Central y Periferico 1Leider Campo100% (1)
- ElectroterapiaDocumento9 páginasElectroterapiaJose David Mendez MarreroAún no hay calificaciones