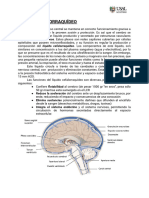
También podría gustarte
- Venciendo Tu AnsiedadDocumento94 páginasVenciendo Tu Ansiedaddiana marcela100% (10)
- Padres Preparados Hijos de CarácterDocumento68 páginasPadres Preparados Hijos de CarácterLorena Figueroa75% (4)
- Test de Ansiedad Generalizada Y VARIOS OTROSDocumento29 páginasTest de Ansiedad Generalizada Y VARIOS OTROSErika Concha Troncoso100% (3)
- Plan de Intervencion en DepresionDocumento9 páginasPlan de Intervencion en DepresionRaquelAún no hay calificaciones
- Resumen Del Sistema NerviosoDocumento5 páginasResumen Del Sistema NerviosoAtenea 03Aún no hay calificaciones
- Depresion de BeckDocumento9 páginasDepresion de BeckNatalie GC0% (1)
- Informe Final Paola Modificado en PDFDocumento126 páginasInforme Final Paola Modificado en PDFkener tejada67% (3)
- Bipolaridad TESISDocumento33 páginasBipolaridad TESISAracely Orellana67% (3)
- Patología de La RetinaDocumento50 páginasPatología de La RetinaRoberto AyalaAún no hay calificaciones
- Fundamentos de Neurociencia ConductualDocumento26 páginasFundamentos de Neurociencia ConductualyiaraAún no hay calificaciones
- Bicarbonato de Sodio en La Cura Del Hongo Del CáncerDocumento19 páginasBicarbonato de Sodio en La Cura Del Hongo Del Cáncerlamente_despierta9562Aún no hay calificaciones
- Primeros Auxilios PsicologicosDocumento35 páginasPrimeros Auxilios PsicologicosKaryMilagros100% (2)
- Apuntes Embriologia Del Sistema NerviosoDocumento15 páginasApuntes Embriologia Del Sistema NerviosoCatalina Soler Lioi100% (1)
- La Salud Desde La Perspectiva IntegralDocumento56 páginasLa Salud Desde La Perspectiva IntegralwaltersingoAún no hay calificaciones
- Apunte de Neurofisiologia Del Dr. Eleri Sworres v1Documento92 páginasApunte de Neurofisiologia Del Dr. Eleri Sworres v1amadoacosta2013100% (1)
- Monografia de Bases Molculares - BiologiaDocumento37 páginasMonografia de Bases Molculares - BiologiaAndres RivasAún no hay calificaciones
- MONOGRAFIA HidrocefaliaDocumento33 páginasMONOGRAFIA HidrocefaliaDavid Llanque60% (5)
- Apunte de Neurofisiologia Del Dr. Eleri Sworres v1 PDFDocumento92 páginasApunte de Neurofisiologia Del Dr. Eleri Sworres v1 PDFJuan RodriguezAún no hay calificaciones
- Resumen Capitulo 6 Apuntes Muy Completos Del LangmanDocumento11 páginasResumen Capitulo 6 Apuntes Muy Completos Del LangmanEMILY CAMILA SARAGURO TENESACAAún no hay calificaciones
- S29 - Informe de SeminarioDocumento11 páginasS29 - Informe de SeminarioJared RequejoAún no hay calificaciones
- Exposicion EmbriologiaDocumento34 páginasExposicion EmbriologiaEstefanni CLAún no hay calificaciones
- Apa AnatomiaDocumento124 páginasApa AnatomiaDiego FigueroaAún no hay calificaciones
- Doctrina Monroe-KellyDocumento6 páginasDoctrina Monroe-KellySebastian CoronadoAún no hay calificaciones
- Organización General - Neurohístología - Estructura Interna y Externa de Los Hemisferios ... Areas Corticales y Neurotransmisores.Documento30 páginasOrganización General - Neurohístología - Estructura Interna y Externa de Los Hemisferios ... Areas Corticales y Neurotransmisores.ADRIANA MUÑOZAún no hay calificaciones
- Usb en NubeDocumento25 páginasUsb en NubeCarlos Tito ObregonAún no hay calificaciones
- L.T.F. Lilia Garcia Razo.: Sistema Nervioso CentralDocumento453 páginasL.T.F. Lilia Garcia Razo.: Sistema Nervioso CentralAdriana De la PeñaAún no hay calificaciones
- NEUROANATOMIADocumento23 páginasNEUROANATOMIAAna GuzmanAún no hay calificaciones
- Bases Neurobiologicas Del Envejecimiento Daniiiii PDFDocumento27 páginasBases Neurobiologicas Del Envejecimiento Daniiiii PDFDaniela AguirreAún no hay calificaciones
- Embriologia Craneofacial DRA RODRIGUEZDocumento8 páginasEmbriologia Craneofacial DRA RODRIGUEZErika Rodriguez MarteAún no hay calificaciones
- Circulación CerebralDocumento3 páginasCirculación CerebralJulieta LinaresAún no hay calificaciones
- Neuroanatomía TextoDocumento154 páginasNeuroanatomía TextoNia AvellanedaAún no hay calificaciones
- Resumen Final de Neuropsicología de FPDocumento26 páginasResumen Final de Neuropsicología de FPFlorpi BronxAún no hay calificaciones
- Nutrición, Protección y Sostén Del SNC - U3 - Parte 1Documento5 páginasNutrición, Protección y Sostén Del SNC - U3 - Parte 1Daniela Figueroa de RayonAún no hay calificaciones
- Semiología Del Paciente en ComaDocumento20 páginasSemiología Del Paciente en ComaRodolfo Leones Castillo100% (1)
- Sistema Nervioso PeriféricoDocumento21 páginasSistema Nervioso Periféricomisionribasguayabal27Aún no hay calificaciones
- Recomendaciones para El Estudio Del Líquido CefalorraquídeoDocumento5 páginasRecomendaciones para El Estudio Del Líquido CefalorraquídeoMaria Belen PeraltaAún no hay calificaciones
- Desarrollo Embrionario Del Sistema Nervioso Central y Órganos de Los Sentidos - RevisiónDocumento9 páginasDesarrollo Embrionario Del Sistema Nervioso Central y Órganos de Los Sentidos - RevisiónCristhiam Anderson B WAún no hay calificaciones
- Embiologia Humana Ii Clase Teorico Practica Del SNDocumento7 páginasEmbiologia Humana Ii Clase Teorico Practica Del SNSantiago AndrésAún no hay calificaciones
- Enfermedades Producidas Por Alteración en La Membrana PlasmáticaDocumento20 páginasEnfermedades Producidas Por Alteración en La Membrana PlasmáticaAngie Nima LadinesAún no hay calificaciones
- Patologías Del SNC Resumen 1Documento8 páginasPatologías Del SNC Resumen 1Marlen BarreraAún no hay calificaciones
- Monografia NeurologiaDocumento25 páginasMonografia NeurologiaCarlos Tito ObregonAún no hay calificaciones
- MONOGRAFIADocumento16 páginasMONOGRAFIAZarella Giuliana Sologuren HumeAún no hay calificaciones
- Monografia Del Sistema Nervioso CentralDocumento31 páginasMonografia Del Sistema Nervioso CentralBill BenassiAún no hay calificaciones
- Liquido CefalorraquideoDocumento11 páginasLiquido CefalorraquideoHomerAún no hay calificaciones
- Informe de Bio Lab 1Documento33 páginasInforme de Bio Lab 1Nazareth de leonAún no hay calificaciones
- Resolver Las Siguientes Preguntas APADocumento3 páginasResolver Las Siguientes Preguntas APAAndyAún no hay calificaciones
- Ictus y HemiplejiaDocumento18 páginasIctus y HemiplejiaJulia JakelineAún no hay calificaciones
- Lectura 3.2Documento13 páginasLectura 3.2MelanyAún no hay calificaciones
- El Líquido Cefalorraquídeo: Ramón J. Ferro ( ) y Rubén L. Makinistian ( )Documento4 páginasEl Líquido Cefalorraquídeo: Ramón J. Ferro ( ) y Rubén L. Makinistian ( )Janeth SanchezAún no hay calificaciones
- Neuroanatomia 1 PDFDocumento79 páginasNeuroanatomia 1 PDFMiguel TorresAún no hay calificaciones
- CAPÍTULO 17 Sistema NerviosoDocumento13 páginasCAPÍTULO 17 Sistema NerviosoTodoMePuedeAlvAún no hay calificaciones
- 2 Mielopatias VascularesDocumento14 páginas2 Mielopatias VascularesOncología Paralelo DAún no hay calificaciones
- Sindromes - Piramidales 02112019Documento22 páginasSindromes - Piramidales 02112019Thiago SantosAún no hay calificaciones
- 3 Carpeta de EmbriologíaDocumento129 páginas3 Carpeta de Embriologíaemelin garciaAún no hay calificaciones
- Primera Parte EmbriologiaDocumento30 páginasPrimera Parte EmbriologiamiguelAún no hay calificaciones
- SN Normal.Documento14 páginasSN Normal.Tomás Gil SantosAún no hay calificaciones
- Nervioso CKQDocumento33 páginasNervioso CKQMatty MarchandAún no hay calificaciones
- Liquido CefalorraquideoDocumento13 páginasLiquido CefalorraquideoACOSTA GARCIA EMILY MARIANAún no hay calificaciones
- Informe Final 12 PDFDocumento47 páginasInforme Final 12 PDFXotchilt Georgina AragonAún no hay calificaciones
- DossierDocumento173 páginasDossierCecilia MAún no hay calificaciones
- PurvecitoDocumento90 páginasPurvecitoBrunna Luiza SimonettiAún no hay calificaciones
- Induccion Molecular Del SNCDocumento37 páginasInduccion Molecular Del SNCAntilla RebazaAún no hay calificaciones
- Unidad 4. Ventrículos Y Líquido Cerebro - Espinal: Mtra. Npsic. Cinthia Alondra Bolaños SánchezDocumento36 páginasUnidad 4. Ventrículos Y Líquido Cerebro - Espinal: Mtra. Npsic. Cinthia Alondra Bolaños SánchezCinthia Alondra Bolaños SánchezAún no hay calificaciones
- 6.19 Suministro de Sangre, Sangre-CerebroDocumento5 páginas6.19 Suministro de Sangre, Sangre-Cerebrojose davidAún no hay calificaciones
- Integracion de Hipertension Endocraneana y Accidente Vascular CerebralDocumento42 páginasIntegracion de Hipertension Endocraneana y Accidente Vascular CerebralARIANA NICOLLE AVILA MENDOZAAún no hay calificaciones
- Neuroanatomia GeronimoDocumento17 páginasNeuroanatomia GeronimoNathalia SilvaAún no hay calificaciones
- Malformaciones Congenitas PDFDocumento152 páginasMalformaciones Congenitas PDFJennyLizbethAún no hay calificaciones
- Catalago Biologia 2Documento16 páginasCatalago Biologia 2rodrigo Acosta mendezAún no hay calificaciones
- Choice Final ForenseDocumento21 páginasChoice Final ForensePili LarramendiAún no hay calificaciones
- Adultez Intermedia y AdultaDocumento15 páginasAdultez Intermedia y AdultapascualAún no hay calificaciones
- Inventario Actualizadotestoteca2017Documento100 páginasInventario Actualizadotestoteca2017Jacqueline Buentello0% (1)
- Bateria de Puebas ClinicasDocumento28 páginasBateria de Puebas Clinicasperroviejo21Aún no hay calificaciones
- Informe de EntrevistaDocumento25 páginasInforme de EntrevistaDaniela Paniagua Castellon0% (1)
- PF IDSR Según VancouverDocumento25 páginasPF IDSR Según VancouverolgaAún no hay calificaciones
- El Punto Fen FuDocumento1 páginaEl Punto Fen Fudianaprieto69Aún no hay calificaciones
- PSICOLOGÍADocumento8 páginasPSICOLOGÍADanny MarrufoAún no hay calificaciones
- Mms Salud ProhibidaDocumento408 páginasMms Salud ProhibidaJc RamAún no hay calificaciones
- Estudio de Caso María Del Pilar Gaviria Jiménez Maestría PsicoterapiaDocumento80 páginasEstudio de Caso María Del Pilar Gaviria Jiménez Maestría PsicoterapiaLaura Ximena HenaoAún no hay calificaciones
- Formato Atencion ClinicaDocumento5 páginasFormato Atencion ClinicaCATALINA OBANDO BENAVIDESAún no hay calificaciones
- Paf CorregidoDocumento27 páginasPaf CorregidoJulissa MegoAún no hay calificaciones
- Informe de MICI IIDocumento27 páginasInforme de MICI IIIvan RodriguezAún no hay calificaciones
- Fichas Textuales ImprimirDocumento3 páginasFichas Textuales ImprimirVanessa Malpartida100% (1)
- Papel Del Psicólogo en Los Centros EducativosDocumento5 páginasPapel Del Psicólogo en Los Centros EducativosMariano DominguezAún no hay calificaciones
- Historia Clínica 2020Documento2 páginasHistoria Clínica 2020BermaryAún no hay calificaciones
- Anteproyecto DefinitivoDocumento36 páginasAnteproyecto DefinitivoEmilia CriptoAún no hay calificaciones
- Unidad de Orientacion EmpresarialDocumento10 páginasUnidad de Orientacion Empresarialenmanuel villalbaAún no hay calificaciones
- 2020 Factores Asociados A Depresión en Enfermeros Del Hospital Emergencias Grau Que Atienden Covid-19Documento71 páginas2020 Factores Asociados A Depresión en Enfermeros Del Hospital Emergencias Grau Que Atienden Covid-19Kehi Meybi Landeo RiveraAún no hay calificaciones
- Fase - Tres - 403008 - 34 U (1) - PSICOPATOLOGIA DE LA ADULTEZ Y LA VEJEZDocumento19 páginasFase - Tres - 403008 - 34 U (1) - PSICOPATOLOGIA DE LA ADULTEZ Y LA VEJEZRodriguez FabianAún no hay calificaciones