También podría gustarte
- Trabajo de InvestigaciónDocumento50 páginasTrabajo de InvestigaciónLadys DunlapAún no hay calificaciones
- 0121 4004 Vitae 25 02 00083Documento9 páginas0121 4004 Vitae 25 02 00083Fito Pratama HelykenAún no hay calificaciones
- SIMILARDocumento17 páginasSIMILARDenis Huanca De La CruzAún no hay calificaciones
- Informe Biotecnologia 1Documento3 páginasInforme Biotecnologia 1Jessica LunaAún no hay calificaciones
- Informe 6 MicroDocumento6 páginasInforme 6 MicroalisonAún no hay calificaciones
- 3 Fábrica-XliDocumento5 páginas3 Fábrica-XliJesus PeraltaAún no hay calificaciones
- Minipreps, Tipos de Biotecnologia y Tipos de PromotoresDocumento12 páginasMinipreps, Tipos de Biotecnologia y Tipos de PromotoresManu RodriguezAún no hay calificaciones
- Tarea 1 Sólido-FluidoDocumento3 páginasTarea 1 Sólido-FluidoAndrea Rojas SuarezAún no hay calificaciones
- Caracterización Bioquímica de AmiJ33, Una Amilasa de Bacillus Amyloliquefaciens Aislada de Suelos CuDocumento22 páginasCaracterización Bioquímica de AmiJ33, Una Amilasa de Bacillus Amyloliquefaciens Aislada de Suelos CuHanylu AvilaAún no hay calificaciones
- Metabolismo Microbiano de Carbohidratos y Lipidos-IV-ciclo-Ing Agroindustrial (Torres Villanueva Mitshell)Documento18 páginasMetabolismo Microbiano de Carbohidratos y Lipidos-IV-ciclo-Ing Agroindustrial (Torres Villanueva Mitshell)Mit TorresAún no hay calificaciones
- Unidad 3 - Fase 4 - Gilberto de Jesús Cano BastoDocumento5 páginasUnidad 3 - Fase 4 - Gilberto de Jesús Cano Bastodayana alvisAún no hay calificaciones
- LEBAME Estabilidad de Este BioproductoDocumento7 páginasLEBAME Estabilidad de Este BioproductoAlejandroAún no hay calificaciones
- Dracaena Trifasciata Estudio FitoquímicoDocumento8 páginasDracaena Trifasciata Estudio FitoquímicoCarlos MendozaAún no hay calificaciones
- Bacterias LacticasDocumento3 páginasBacterias LacticasLeonardo SilvaAún no hay calificaciones
- UDLA-EC-TIB-2019-19 Bacillus Subtilis Atomizacion y LiofilizacionDocumento60 páginasUDLA-EC-TIB-2019-19 Bacillus Subtilis Atomizacion y LiofilizacionOscar CotaAún no hay calificaciones
- Entregable 1-Bases MolecularesDocumento7 páginasEntregable 1-Bases MolecularesAna Paola Morales MendozaAún no hay calificaciones
- Uso de Nanoparticulas de QuitosanoDocumento12 páginasUso de Nanoparticulas de QuitosanoAbdias Machacuay PaitaAún no hay calificaciones
- Coleccion y Conservacion de MicroorganisDocumento8 páginasColeccion y Conservacion de MicroorganisMARIA DE LA PAZ GRAJEDA PINEDAAún no hay calificaciones
- Kluyveromyces MarxianusDocumento7 páginasKluyveromyces MarxianusAgentesteveAún no hay calificaciones
- Borrador e Info Del ProyectoDocumento3 páginasBorrador e Info Del ProyectoElizabeth Natali Sanchez RamirezAún no hay calificaciones
- Aplicaciones de La BacteriologiaDocumento4 páginasAplicaciones de La BacteriologiaEduardo Rodriguez0% (1)
- Proyecto Equipo 1 - Regulacion Metabolica de L. DelbrueckiiDocumento22 páginasProyecto Equipo 1 - Regulacion Metabolica de L. DelbrueckiiJorge OlmedoAún no hay calificaciones
- Trabajo II de Osbel Núñez, Atagua (1) - 0410113159Documento11 páginasTrabajo II de Osbel Núñez, Atagua (1) - 0410113159Joyssi LopezAún no hay calificaciones
- Informe de Laboratorio - Extracción de Adn BacterianoDocumento6 páginasInforme de Laboratorio - Extracción de Adn BacterianoAndres DavidAún no hay calificaciones
- Investigación Bibliográfica Grupo 3Documento14 páginasInvestigación Bibliográfica Grupo 3Iris109Aún no hay calificaciones
- Traduccion Paper 2Documento11 páginasTraduccion Paper 2Kyle Valentine CollenAún no hay calificaciones
- 2021 RMIQ v202 Pag711-723 JCR FI1068 220517 193442 220608 100002Documento14 páginas2021 RMIQ v202 Pag711-723 JCR FI1068 220517 193442 220608 10000215041168Aún no hay calificaciones
- Resumen Bacteriocinas en Productos CarnicosDocumento3 páginasResumen Bacteriocinas en Productos CarnicosandresitotsAún no hay calificaciones
- Modelameinto y Re-Ingenieria de EC 3.4.21.62 - Subtilisina de Bacillus SubtilisDocumento37 páginasModelameinto y Re-Ingenieria de EC 3.4.21.62 - Subtilisina de Bacillus SubtilisCarlos Humberto Gomez RodriguezAún no hay calificaciones
- Graficas DPPH Abst ESDocumento15 páginasGraficas DPPH Abst ESLourdes VictoriaAún no hay calificaciones
- Informen°8 Lab BioprocesosDocumento8 páginasInformen°8 Lab BioprocesosMARIA DEL PILAR VILCHEZ MOLINAAún no hay calificaciones
- Gamba2021 Article ChangesInMicrobiotaDuringMultiDocumento8 páginasGamba2021 Article ChangesInMicrobiotaDuringMultilaura Montoya ZapataAún no hay calificaciones
- Informe No. 1 - BioprocesosDocumento5 páginasInforme No. 1 - BioprocesosAndrésFelipeQuirogaMirandaAún no hay calificaciones
- Compuestos Con Posible Actividad Fotoprotectora A Partir de Extractos de Macroalgas Del Caribe ColombianoDocumento236 páginasCompuestos Con Posible Actividad Fotoprotectora A Partir de Extractos de Macroalgas Del Caribe ColombianoMaria OviedoAún no hay calificaciones
- Síntesis de N-Glucopéptidos Derivados de La Lfcinb Evaluación de Su Actividad Antibacteriana AntifúngicaDocumento141 páginasSíntesis de N-Glucopéptidos Derivados de La Lfcinb Evaluación de Su Actividad Antibacteriana AntifúngicaAngie RozoAún no hay calificaciones
- Clonación, Expresión, Caracterización y Modelado deDocumento13 páginasClonación, Expresión, Caracterización y Modelado deLucia HernándezAún no hay calificaciones
- RESUMEN ARTICULOS PBL TermoDocumento2 páginasRESUMEN ARTICULOS PBL TermoSantiago Tamayo MarinAún no hay calificaciones
- Congreso Latinoamericano de Aguacate PDFDocumento8 páginasCongreso Latinoamericano de Aguacate PDFESTEBAN JARAMILLO JARAMILLOAún no hay calificaciones
- Polímeros de CarbohidratosDocumento15 páginasPolímeros de CarbohidratosMonica ZuletaAún no hay calificaciones
- Villalva Et Al., 2018.en - EsDocumento33 páginasVillalva Et Al., 2018.en - EskatherinetAún no hay calificaciones
- Evaluación de La Actividad Antioxidante de Extractos de Flavonoides de Cáscara de Naranja en El Aceite de Soja DesodorizadoDocumento8 páginasEvaluación de La Actividad Antioxidante de Extractos de Flavonoides de Cáscara de Naranja en El Aceite de Soja DesodorizadoRaul Toledo RodriguezAún no hay calificaciones
- Informe 6 Micro Ind (Esterilidad Comercial)Documento7 páginasInforme 6 Micro Ind (Esterilidad Comercial)Daniel Giraldo0% (1)
- Tp-N12-Microbiologia BAL Y BACDocumento4 páginasTp-N12-Microbiologia BAL Y BACSusana BenittiAún no hay calificaciones
- UAMI19140Documento176 páginasUAMI19140Yisela EscobarAún no hay calificaciones
- Mantenimiento y Conservación de MicroorganismosDocumento7 páginasMantenimiento y Conservación de Microorganismosjorge eduardoAún no hay calificaciones
- Antibacterial Activity of The Hidroalcoholic Extract Croton Draco On Bacteria of Sanitary ImportanceDocumento10 páginasAntibacterial Activity of The Hidroalcoholic Extract Croton Draco On Bacteria of Sanitary ImportanceESMERALDA TOROAún no hay calificaciones
- UntitledDocumento9 páginasUntitledValeria Carranza HerreraAún no hay calificaciones
- Ficha Técnica ATCCDocumento6 páginasFicha Técnica ATCCDiego Gpe Ac AcAún no hay calificaciones
- ARTÍCULO Actividades Antioxidantes e Inducción de Apoptosis Por Extractos de Metanol de Aguacate ESPAÑOLDocumento7 páginasARTÍCULO Actividades Antioxidantes e Inducción de Apoptosis Por Extractos de Metanol de Aguacate ESPAÑOLMónica MartínezAún no hay calificaciones
- Informe N 8 Metabolismo MicrobianoDocumento7 páginasInforme N 8 Metabolismo MicrobianoelianAún no hay calificaciones
- Bioquimica BacterianaDocumento15 páginasBioquimica BacterianaRenzoDanovicAún no hay calificaciones
- Reporte 5 - EQ1 - 2BM1 - LTMDocumento20 páginasReporte 5 - EQ1 - 2BM1 - LTMMAURICIO SOSA GARCIAAún no hay calificaciones
- Desinfectante de AlimentosDocumento6 páginasDesinfectante de Alimentosjacks_danilAún no hay calificaciones
- Informe #2 BioquímicaDocumento6 páginasInforme #2 Bioquímicalaurabargo2004Aún no hay calificaciones
- Tesis Deternacion de Lactobacilus en Arina de LinazaDocumento91 páginasTesis Deternacion de Lactobacilus en Arina de LinazaRonald PalominoAún no hay calificaciones
- BioingenieríaDocumento8 páginasBioingenieríajesusAún no hay calificaciones
- 35035Documento114 páginas35035Viviana CastilloAún no hay calificaciones
- PRÁCTICA FINAL DE LABORATORIO MicrobiologìaDocumento29 páginasPRÁCTICA FINAL DE LABORATORIO MicrobiologìaDAYANARA ELIANA YUPTON CALDERONAún no hay calificaciones
- Avances de la Biotecnología y su impacto en la agricultura latinoamericanaDe EverandAvances de la Biotecnología y su impacto en la agricultura latinoamericanaAún no hay calificaciones
- Técnicas ómicas aplicadas al estudio de la microbiotaDe EverandTécnicas ómicas aplicadas al estudio de la microbiotaAún no hay calificaciones
- Analisis UTFSMfisicoquimicDocumento65 páginasAnalisis UTFSMfisicoquimicCristian VillarAún no hay calificaciones
- controlador cloroDocumento129 páginascontrolador cloroCristian VillarAún no hay calificaciones
- Vigilancia-Nuevas Variantes CovidDocumento43 páginasVigilancia-Nuevas Variantes CovidCristian VillarAún no hay calificaciones
- Componenets Oi TDocumento11 páginasComponenets Oi TCristian VillarAún no hay calificaciones
- I A+bioinformaticaDocumento13 páginasI A+bioinformaticaCristian VillarAún no hay calificaciones
- Ingenieria MetabolicaDocumento3 páginasIngenieria MetabolicaCristian VillarAún no hay calificaciones
- Analisis de La Mutacion d614g - Version Final - 22 Enero 2021 - H - 9 31 MDocumento6 páginasAnalisis de La Mutacion d614g - Version Final - 22 Enero 2021 - H - 9 31 MCristian VillarAún no hay calificaciones
- f19 Consultia ChemoDocumento39 páginasf19 Consultia ChemoCristian VillarAún no hay calificaciones
- Quib0108 Gestion Y Control de Planta Quimica OnlineDocumento42 páginasQuib0108 Gestion Y Control de Planta Quimica OnlineCristian VillarAún no hay calificaciones
- PDB OlivaresHernándezA CáncerDocumento211 páginasPDB OlivaresHernándezA CáncerCristian VillarAún no hay calificaciones
- TFG - Diaz Lozano, BeatrizDocumento269 páginasTFG - Diaz Lozano, BeatrizCristian VillarAún no hay calificaciones
- Syllabus Contenidos MateriasDocumento4 páginasSyllabus Contenidos MateriasCristian VillarAún no hay calificaciones
- 181 Revista de La Sociedad Argentina de GeneticaDocumento278 páginas181 Revista de La Sociedad Argentina de GeneticaCristian VillarAún no hay calificaciones
- Juzgado de Letras y Garantía de Lebu.: Andrea Valeria Rodríguez FerradaDocumento1 páginaJuzgado de Letras y Garantía de Lebu.: Andrea Valeria Rodríguez FerradaCristian VillarAún no hay calificaciones
- Guia Didactica Curso Consultoria Sap PP-PMDocumento7 páginasGuia Didactica Curso Consultoria Sap PP-PMCristian VillarAún no hay calificaciones
- Manual de Usuario de Caiba de BioseguridadDocumento51 páginasManual de Usuario de Caiba de BioseguridadEstefanía GuevaraAún no hay calificaciones
- 0041 9095 Unmed 61 02 00049Documento15 páginas0041 9095 Unmed 61 02 00049Joe ArajAún no hay calificaciones
- Tema 11 Investigacion Operativa Con RDocumento10 páginasTema 11 Investigacion Operativa Con RTito BarrionuevoAún no hay calificaciones
- 0041 9095 Unmed 61 02 00049Documento15 páginas0041 9095 Unmed 61 02 00049Joe ArajAún no hay calificaciones
- Predicción en El Diagnóstico de Tumores de Cáncer de Mama Empleando Métodos de ClasificaciónDocumento9 páginasPredicción en El Diagnóstico de Tumores de Cáncer de Mama Empleando Métodos de ClasificaciónCristian VillarAún no hay calificaciones
- Analiss Biomecanico Del Futbolista en El ChuteDocumento5 páginasAnaliss Biomecanico Del Futbolista en El ChuteRamiro RoblesAún no hay calificaciones
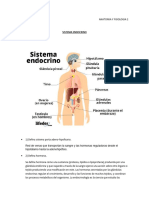
- Sistema EndocrinoDocumento4 páginasSistema Endocrinofernanda carabajalAún no hay calificaciones
- FISIOLOGIA Del Sistema UrinarioDocumento36 páginasFISIOLOGIA Del Sistema UrinarioMayori Lisseth Ruiz AguirreAún no hay calificaciones
- Glosario de Terminos NeurologicosDocumento22 páginasGlosario de Terminos NeurologicosGuillermo MunozAún no hay calificaciones
- Resultados SaludDignaDocumento2 páginasResultados SaludDignaEsmeralda Meza Felix100% (1)
- Maniobras SemiologicasDocumento94 páginasManiobras SemiologicasANGULO REAS VICTOR ALEJANDROAún no hay calificaciones
- Riñones en Especies AnimalesDocumento3 páginasRiñones en Especies AnimalesFernando TauferAún no hay calificaciones
- Anatomia Del PeritoneoDocumento26 páginasAnatomia Del PeritoneoCarolina Herrera100% (1)
- Examen de BiologiaDocumento3 páginasExamen de BiologiaSteven Esteve100% (1)
- Reporte 5 Quimiotaxis y FagocitosisDocumento4 páginasReporte 5 Quimiotaxis y FagocitosisDIANA REYES ESCOBEDOAún no hay calificaciones
- Resumen Capitulo 75Documento8 páginasResumen Capitulo 75Gabo GutierrezAún no hay calificaciones
- Capitulo Ii RiñonDocumento36 páginasCapitulo Ii Riñonadrianamargarita0007.amooAún no hay calificaciones
- Mallas Bachillerato Ciencias Biomedicas-Admision 2021Documento1 páginaMallas Bachillerato Ciencias Biomedicas-Admision 2021diegoaf07Aún no hay calificaciones
- EMBRIOLOGÍA CAVIDADES CORPORALES y RESPIRATORIODocumento13 páginasEMBRIOLOGÍA CAVIDADES CORPORALES y RESPIRATORIOannabella del vecchioAún no hay calificaciones
- Sistema NerviosoDocumento10 páginasSistema NerviosoValentina uwuAún no hay calificaciones
- Guia de Laboratorio Teórica Sistema Muscular PDFDocumento7 páginasGuia de Laboratorio Teórica Sistema Muscular PDFjoselynAún no hay calificaciones
- Manual Electromiografia (Osteomuscular)Documento11 páginasManual Electromiografia (Osteomuscular)Javier JimenezAún no hay calificaciones
- La Biología Como CienciaDocumento19 páginasLa Biología Como CienciaHenry ChumpiAún no hay calificaciones
- Documento Formateado Con Normas APA Sistema RespiratorioDocumento23 páginasDocumento Formateado Con Normas APA Sistema RespiratoriomoranchullymoisesdavidAún no hay calificaciones
- Lectura Complementaria 7 - Respiración CelularDocumento9 páginasLectura Complementaria 7 - Respiración CelularDanna Alecksei Salvador ArizaAún no hay calificaciones
- Fisiología de La Nutrición AnimalDocumento16 páginasFisiología de La Nutrición Animalconstanza salgadoAún no hay calificaciones
- Lupus PatologiaDocumento16 páginasLupus Patologiaswlima-esAún no hay calificaciones
- HTA Adulto Mayor LDocumento35 páginasHTA Adulto Mayor LJess TiradoAún no hay calificaciones
- Patrón Cara y CuelloDocumento6 páginasPatrón Cara y CuelloTatiana MartinezAún no hay calificaciones
- Itsmo de Las FaucesDocumento8 páginasItsmo de Las FaucesEleanor JamesAún no hay calificaciones
- Examen de Medio CursoDocumento7 páginasExamen de Medio CursoMarco MarinAún no hay calificaciones
- Act. 6 Crecimiento Fisico y DesarrolloDocumento17 páginasAct. 6 Crecimiento Fisico y DesarrolloAbigail CruzAún no hay calificaciones
- Movimientos Dentarios en OrtodonciaDocumento23 páginasMovimientos Dentarios en OrtodonciaAranzazu Manjarrez MedellinAún no hay calificaciones
- Rood, Bobath, FNP, Medek, VojtaDocumento16 páginasRood, Bobath, FNP, Medek, VojtaFranyolis Nessi100% (1)
- Ciencia y Ambiente 23.03.2022 MDocumento3 páginasCiencia y Ambiente 23.03.2022 MVladimir Medina QuizaAún no hay calificaciones