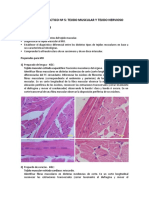
También podría gustarte
- Guia Extraordinario 2022-2 BBCDocumento14 páginasGuia Extraordinario 2022-2 BBCERIKA PAREDESAún no hay calificaciones
- HG 5Documento9 páginasHG 5CrockadockaAún no hay calificaciones
- Anatomia de La Neurona, Cuerpo Celular y Prolongaciones, MielinaDocumento10 páginasAnatomia de La Neurona, Cuerpo Celular y Prolongaciones, MielinaidnyzAún no hay calificaciones
- 28016-Texto Del Artã Culo-43377-1-10-20191010Documento9 páginas28016-Texto Del Artã Culo-43377-1-10-20191010MARCO ANTONIO TAPIA MALDONADOAún no hay calificaciones
- Clase 02 - GlíaDocumento45 páginasClase 02 - GlíaPaola Huaco QuispeAún no hay calificaciones
- Informe Bio Semana 10Documento4 páginasInforme Bio Semana 10Mariana Elizabeth Loja LuceroAún no hay calificaciones
- Práctica Del Encéfalo (Recuperado Automáticamente)Documento6 páginasPráctica Del Encéfalo (Recuperado Automáticamente)Daniela CastilloAún no hay calificaciones
- Celulas Nerviosas y ConductaDocumento22 páginasCelulas Nerviosas y ConductaIsabel Cristina Orozco Bedoya100% (1)
- Guía 1 Adn XenopusDocumento2 páginasGuía 1 Adn XenopusUrrutia O´rian Araceli IsabelAún no hay calificaciones
- Seminario 9 Tejido Nervioso 2007modif Javier2016Documento87 páginasSeminario 9 Tejido Nervioso 2007modif Javier2016Sha KnidAún no hay calificaciones
- Clase Ictiologia Neotropical 2022Documento62 páginasClase Ictiologia Neotropical 2022AcuariofiliadcAún no hay calificaciones
- A24 N87 RegenerationDocumento17 páginasA24 N87 RegenerationCristianAún no hay calificaciones
- Anatomía e Histología de La RetinaDocumento10 páginasAnatomía e Histología de La RetinaReiniel Díaz Pérez100% (1)
- 2 - Principios Del Funcionamiento Del Sistema NerviosoDocumento42 páginas2 - Principios Del Funcionamiento Del Sistema NerviosoDiana HaAún no hay calificaciones
- Cap 6Documento30 páginasCap 6usuario desconocidoAún no hay calificaciones
- Protocolo FisiologiaDocumento15 páginasProtocolo FisiologiaDioneone Manzana PoepleAún no hay calificaciones
- Celulas CiliadasDocumento18 páginasCelulas CiliadasPedro M Malave VAún no hay calificaciones
- Neurobiología de La NeuronaDocumento7 páginasNeurobiología de La NeuronaLeandra MartinelliAún no hay calificaciones
- Guia de Lab Morfo Locomotor 2020 1Documento72 páginasGuia de Lab Morfo Locomotor 2020 1Leydi BravoAún no hay calificaciones
- Ramón y Cajal y La Neurociencia Del Siglo XXI RESUMENDocumento2 páginasRamón y Cajal y La Neurociencia Del Siglo XXI RESUMENJoel CoriaAún no hay calificaciones
- Capítulo de Atlas Citología e Histología 2005 Wolfgang PDFDocumento12 páginasCapítulo de Atlas Citología e Histología 2005 Wolfgang PDFHannie BertalotAún no hay calificaciones
- Neuro Retina 2452 22-2Documento59 páginasNeuro Retina 2452 22-2Montserrat FuentesAún no hay calificaciones
- Metodos y Tecnica en La FisiologiaDocumento7 páginasMetodos y Tecnica en La FisiologiaAlely Moya RodriguezAún no hay calificaciones
- Neurona y NeurogliaDocumento27 páginasNeurona y NeurogliaTory MendozaAún no hay calificaciones
- 2019 RMSAnatoma Bsica Ganglios BasalesDocumento7 páginas2019 RMSAnatoma Bsica Ganglios BasalesLuz ChambiAún no hay calificaciones
- Ferrus, A Rev NeurolDocumento7 páginasFerrus, A Rev NeurolAna Isabel CastañedaAún no hay calificaciones
- 401-Article Text-1494-1-10-20200915Documento6 páginas401-Article Text-1494-1-10-20200915santiago santos baenaAún no hay calificaciones
- Via AuditivaDocumento11 páginasVia AuditivaPaula O.R100% (1)
- Práctica Tinsión de NisslDocumento8 páginasPráctica Tinsión de NisslKaren RodriguezAún no hay calificaciones
- Bioquímica de La Visión IIDocumento11 páginasBioquímica de La Visión IISergio Coronado RojasAún no hay calificaciones
- BYNC II NeuronaDocumento25 páginasBYNC II NeuronaPaula PajónAún no hay calificaciones
- Guia Porifera LabDocumento3 páginasGuia Porifera LabCamilo Andres Espinosa RomeroAún no hay calificaciones
- Repaso Simulagro de Tres Editores.Documento17 páginasRepaso Simulagro de Tres Editores.Valentina MuñozAún no hay calificaciones
- Primer Examen.: A) Cierto B) FalsoDocumento6 páginasPrimer Examen.: A) Cierto B) FalsoJesús Torres MayaAún no hay calificaciones
- Clase 1 - Introduccion A La HistologíaDocumento42 páginasClase 1 - Introduccion A La HistologíaGAJARDO MUNOZ FLORENCIA ANTONAún no hay calificaciones
- Lab 9 MicroDocumento60 páginasLab 9 MicroTatiana HernándezAún no hay calificaciones
- El Cerebro AveriadoDocumento8 páginasEl Cerebro AveriadoSmallReadingRat50% (2)
- Las Anomalias de La Actividad Cerebelosa de Los AutistasDocumento11 páginasLas Anomalias de La Actividad Cerebelosa de Los AutistasNoé Motos GonzálezAún no hay calificaciones
- Metodos en Neurociencias CognitivasDocumento11 páginasMetodos en Neurociencias CognitivasMatías Quintanilla GonzálezAún no hay calificaciones
- Daneri Muzio - Estudio de Las Bases Neurobiológicas de Algunas Patologías Cerebrales Utilizando Técnicas de Diagnóstico Por ImágenesDocumento14 páginasDaneri Muzio - Estudio de Las Bases Neurobiológicas de Algunas Patologías Cerebrales Utilizando Técnicas de Diagnóstico Por ImágenesMaría Luz0% (1)
- Taller de Histología 2 SemestreDocumento21 páginasTaller de Histología 2 SemestreSantiago PalaciosAún no hay calificaciones
- Proyecto Ojos Jovenes InvestigacionDocumento16 páginasProyecto Ojos Jovenes InvestigacionMayte IbanezAún no hay calificaciones
- El OjoDocumento8 páginasEl OjoUrania Serena Moreno GranadosAún no hay calificaciones
- Técnicas de Investigación en NeurofisiologiaDocumento2 páginasTécnicas de Investigación en NeurofisiologiaJaime Jose Arias CongrainsAún no hay calificaciones
- El CerebeloDocumento7 páginasEl Cerebeloquispearapaabraham452Aún no hay calificaciones
- Complementar - Neuronal Loss in Functional Zones - En.esDocumento10 páginasComplementar - Neuronal Loss in Functional Zones - En.esSantiago AlzateAún no hay calificaciones
- Espinas y FilopodiosDocumento12 páginasEspinas y FilopodiosIvan AndresAún no hay calificaciones
- Brodman AreasDocumento36 páginasBrodman Areasabi100% (1)
- 2010 Distribución Intraneural Del Nervio CubitalDocumento4 páginas2010 Distribución Intraneural Del Nervio CubitalDavid Alejandro Cavieres AcuñaAún no hay calificaciones
- Transduccion VisualDocumento7 páginasTransduccion Visualpp0% (1)
- Histología Del Sistema Nervioso CentralDocumento25 páginasHistología Del Sistema Nervioso CentralCesar Garcia NKAún no hay calificaciones
- Lab 9 Manual MicroanatomiaDocumento20 páginasLab 9 Manual MicroanatomiageovannyAún no hay calificaciones
- Bases Biológicas Del ComportamientoDocumento4 páginasBases Biológicas Del ComportamientoROBIN BRANDON ROJAS ALAVAAún no hay calificaciones
- Bases Biológicas Del ComportamientoDocumento4 páginasBases Biológicas Del ComportamientoROBIN BRANDON ROJAS ALAVAAún no hay calificaciones
- Resultados MicrosDocumento8 páginasResultados MicrosAlejandraAún no hay calificaciones
- Espina BífidaDocumento16 páginasEspina BífidaNathalia GaitánAún no hay calificaciones
- Tabajo IndividualDocumento22 páginasTabajo IndividualSebastian Enriquez MancillaAún no hay calificaciones
- Neuroanatomia Patologica Neuropilo Protocolo de Autopsia Micro Agosto 25 2022Documento2 páginasNeuroanatomia Patologica Neuropilo Protocolo de Autopsia Micro Agosto 25 2022nievesrionegroAún no hay calificaciones
- Fragmento Del Capítulo 2 de KandelDocumento5 páginasFragmento Del Capítulo 2 de KandelJuli RasguidoAún no hay calificaciones
- CRISPR Cas-9: ¿Cómo Una Herramienta Genética Puede Ser La Razón de Disputa Global?Documento24 páginasCRISPR Cas-9: ¿Cómo Una Herramienta Genética Puede Ser La Razón de Disputa Global?Nicolle CárdenasAún no hay calificaciones
- Harina de Bagazo de UvaDocumento18 páginasHarina de Bagazo de UvaJoseAún no hay calificaciones
- Examenes de Hematologia Clinica - UnprgDocumento12 páginasExamenes de Hematologia Clinica - UnprgEsther EstAún no hay calificaciones
- Parcial 2Documento88 páginasParcial 2Dianaa Isabella FernandezAún no hay calificaciones
- Areas Deforestadas HumedalesDocumento74 páginasAreas Deforestadas HumedalesFreddy Arcos AlarcónAún no hay calificaciones
- Universidad Mayor de San Andres Carrera de Ingenieria AgronomicaDocumento109 páginasUniversidad Mayor de San Andres Carrera de Ingenieria AgronomicaAlexander CHAún no hay calificaciones
- Clase 10 Plasticidad Del Cerebro AdolescenteDocumento2 páginasClase 10 Plasticidad Del Cerebro AdolescenteCatalina Gómez PasettiAún no hay calificaciones
- Lesiones CervicalesDocumento6 páginasLesiones CervicalesJuan Carlos GutiérrezAún no hay calificaciones
- Melanoma VulvarDocumento58 páginasMelanoma VulvarBenjamin Gerardo Delgado MoralesAún no hay calificaciones
- SISE Bioquimica 1Documento25 páginasSISE Bioquimica 1Frefire MaxAún no hay calificaciones
- Tinnitus y Otoemisiones AcústicasDocumento15 páginasTinnitus y Otoemisiones AcústicasDanielAún no hay calificaciones
- "La Evolución de La Normalidad en La Niñez." Anna FreudDocumento10 páginas"La Evolución de La Normalidad en La Niñez." Anna FreudDiego Valentina Mansilla SilveiraAún no hay calificaciones
- Texto. El Gato. Práctico. 1er Año PrimariaDocumento1 páginaTexto. El Gato. Práctico. 1er Año PrimariaPatri SaucedoAún no hay calificaciones
- Catalogo de Variedades de Papa Nativa en Mérida, VenezuelaDocumento100 páginasCatalogo de Variedades de Papa Nativa en Mérida, VenezuelaJorge Luis Alonso G.100% (4)
- Prevencion de Lesiones OsteomuscularesDocumento11 páginasPrevencion de Lesiones OsteomuscularesAlvaro VianaAún no hay calificaciones
- Sistema HemáticoDocumento22 páginasSistema HemáticoAlejandra PaolaAún no hay calificaciones
- SAEZ, C. Fertilidad de Los Suelos de La Región de Magallanes (1995)Documento32 páginasSAEZ, C. Fertilidad de Los Suelos de La Región de Magallanes (1995)agropatagoniaAún no hay calificaciones
- Cohen Dorothy - Aspectos Del Desarrollo de Los Niños de 5 Años..Documento10 páginasCohen Dorothy - Aspectos Del Desarrollo de Los Niños de 5 Años..Janeth Garcia Sosa100% (1)
- Documento Plan de Manejo PN Toro Toro 2013-2022Documento238 páginasDocumento Plan de Manejo PN Toro Toro 2013-2022Carola Miranda100% (2)
- Ensayo Teroria Del CreasionismoDocumento2 páginasEnsayo Teroria Del CreasionismoLuis Alberto GAún no hay calificaciones
- Jornadas Micologicas SIERRA NEVADADocumento9 páginasJornadas Micologicas SIERRA NEVADAapi-19937287Aún no hay calificaciones
- Propiedades Intensivas y ExtensivasDocumento7 páginasPropiedades Intensivas y ExtensivasGeorge AdivinaAún no hay calificaciones
- NUTRICIONDocumento2 páginasNUTRICIONCarlosAún no hay calificaciones
- Nivel 1.1-Caracteristicas Del Acondicionamiento FisicoDocumento14 páginasNivel 1.1-Caracteristicas Del Acondicionamiento FisicoMiguel SanchezAún no hay calificaciones
- Sistema RadicularDocumento20 páginasSistema RadicularyesicolAún no hay calificaciones
- Cuestionario de Estados Reiterados de Frustración .Documento7 páginasCuestionario de Estados Reiterados de Frustración .karina paterninaAún no hay calificaciones
- Semana 4 - Grado Cuarto - Guias Aprender en Casa - KARENTBDocumento3 páginasSemana 4 - Grado Cuarto - Guias Aprender en Casa - KARENTBJhonatan Esteban Melgarejo CarreñoAún no hay calificaciones
- Wuolah Free TEMA 1Documento8 páginasWuolah Free TEMA 1MARTA WARAún no hay calificaciones
- Sindrome FebrilDocumento3 páginasSindrome FebrilAndrés OcampoAún no hay calificaciones
- Guías Del Laboratorio de BiologíaDocumento70 páginasGuías Del Laboratorio de BiologíaSara GonzálezAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- 7 Pasos Para Superar Una Crisis: Cómo atravesar un duelo o una pérdida y salir fortalecidoDe Everand7 Pasos Para Superar Una Crisis: Cómo atravesar un duelo o una pérdida y salir fortalecidoCalificación: 5 de 5 estrellas5/5 (5)
- Las Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishDe EverandLas Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Curación a través de Un Curso de MilagrosDe EverandCuración a través de Un Curso de MilagrosCalificación: 4.5 de 5 estrellas4.5/5 (4)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Los 12 chakras: Desbloquea tus dones espiritualesDe EverandLos 12 chakras: Desbloquea tus dones espiritualesCalificación: 4 de 5 estrellas4/5 (14)
- Entrenamiento Científico con pesas: Fitness InteligenteDe EverandEntrenamiento Científico con pesas: Fitness InteligenteCalificación: 5 de 5 estrellas5/5 (3)
- Método Montessori para bebés de 0 a 3 años. Guía práctica y útil para criar niños tranquilos, competentes y segurosDe EverandMétodo Montessori para bebés de 0 a 3 años. Guía práctica y útil para criar niños tranquilos, competentes y segurosAún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Código Mujer: Womancode: Optimiza tu ciclo, aumenta tu fertilidad, potencia tu deseo sexual y conviértete en una fuente de energíaDe EverandCódigo Mujer: Womancode: Optimiza tu ciclo, aumenta tu fertilidad, potencia tu deseo sexual y conviértete en una fuente de energíaCalificación: 5 de 5 estrellas5/5 (2)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- La curación espontánea de las creencias: Cómo librarse de los falsos límitesDe EverandLa curación espontánea de las creencias: Cómo librarse de los falsos límitesCalificación: 4.5 de 5 estrellas4.5/5 (22)
- Cómo dejar el patrón de obesidad con el trabajo de constelaciones sistémicasDe EverandCómo dejar el patrón de obesidad con el trabajo de constelaciones sistémicasCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Dejar ir. El camino a la liberaciónDe EverandDejar ir. El camino a la liberaciónCalificación: 4.5 de 5 estrellas4.5/5 (22)
- Un intestino feliz. Cómo la microbiota mejora tu salud mental y te ayuda a manejar las emocionesDe EverandUn intestino feliz. Cómo la microbiota mejora tu salud mental y te ayuda a manejar las emocionesCalificación: 5 de 5 estrellas5/5 (4)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Cardiología y enfermedades cardiovascularesDe EverandCardiología y enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Libro de cocina completo para diabéticos en español/ Diabetic cookbook in spanishDe EverandLibro de cocina completo para diabéticos en español/ Diabetic cookbook in spanishCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Shiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasDe EverandShiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasAún no hay calificaciones
- La vida antes de nacer: Viaje al origen de nuestra historia personalDe EverandLa vida antes de nacer: Viaje al origen de nuestra historia personalCalificación: 5 de 5 estrellas5/5 (3)
- En sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésDe EverandEn sintonía con tu ciclo femenino: FLO. Aprende a sincronizarte con tu bioquímica para dar rienda suelta a tu creatividad, mejorar tu vida y hacer más con menos estrésCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Joven a cualquier edad. El método definitivo para una vida larga, saludable y felizDe EverandJoven a cualquier edad. El método definitivo para una vida larga, saludable y felizAún no hay calificaciones
- Nuevo manual de Reflexología: El método más completo y actual sobre las técnicas, la práctica y la teoría de la ciencia reflexológicaDe EverandNuevo manual de Reflexología: El método más completo y actual sobre las técnicas, la práctica y la teoría de la ciencia reflexológicaCalificación: 4.5 de 5 estrellas4.5/5 (16)
- Postres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaDe EverandPostres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaCalificación: 5 de 5 estrellas5/5 (17)