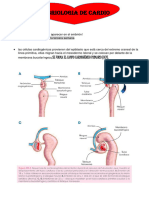
También podría gustarte
- Desarrollo Del Sistema CardiovascularDocumento11 páginasDesarrollo Del Sistema CardiovascularAlicia Hernández PérezAún no hay calificaciones
- Desarrollo Embrionario Del Sistema CardiovascularDocumento17 páginasDesarrollo Embrionario Del Sistema CardiovascularfavalosconejerosAún no hay calificaciones
- HistoembriologíaDocumento24 páginasHistoembriologíaMartin LudeñaAún no hay calificaciones
- Sistema CirculatorioDocumento12 páginasSistema CirculatorioJessica Andrea Alamilla GarcíaAún no hay calificaciones
- Sistema Cardiovascular embriologíaDocumento4 páginasSistema Cardiovascular embriologíaLuna JoyaAún no hay calificaciones
- Embriología - Desarrollo Embriológico Del Sistema Cardiovascular - Corazón y Grandes VasosDocumento41 páginasEmbriología - Desarrollo Embriológico Del Sistema Cardiovascular - Corazón y Grandes VasosDoris Sarai Fernández VelaAún no hay calificaciones
- Resumen Embriologia Del Corazon PDFDocumento6 páginasResumen Embriologia Del Corazon PDFAlfredo LlerenaAún no hay calificaciones
- Sem 14-Campos Cardiogénicos y Formación de La Asa CardiacaDocumento8 páginasSem 14-Campos Cardiogénicos y Formación de La Asa CardiacaEduar OspinaAún no hay calificaciones
- Anatomía Mod 4Documento49 páginasAnatomía Mod 4javiergarridoabAún no hay calificaciones
- Sistema CardiovascularDocumento12 páginasSistema CardiovascularCristina VelázquezAún no hay calificaciones
- CardiogénesisDocumento5 páginasCardiogénesisNath Rangel (Naty)Aún no hay calificaciones
- 07-UNIDAD III Embriologia Del CorazonDocumento56 páginas07-UNIDAD III Embriologia Del CorazonCesia J. Darce100% (1)
- Resumen Sistema CardiovascularDocumento14 páginasResumen Sistema CardiovascularVanessa CastañedaAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento25 páginasEmbriologia Del Sistema CardiovascularLeyla PerezAún no hay calificaciones
- Sistema CardiovascularDocumento98 páginasSistema CardiovascularIncy LopezAún no hay calificaciones
- Asa CardiacaDocumento2 páginasAsa CardiacaBRYAN HERRERAAún no hay calificaciones
- CARDIOVASCULARDocumento4 páginasCARDIOVASCULARJhoswil DamianAún no hay calificaciones
- 4 Resumen Del Sistema CardiovascularDocumento4 páginas4 Resumen Del Sistema CardiovascularosnelAún no hay calificaciones
- SISTEMA CARDIOVASCULAR UwUDocumento13 páginasSISTEMA CARDIOVASCULAR UwUPaola AvilaAún no hay calificaciones
- Desarrollo Del Sistema CardiovascularDocumento12 páginasDesarrollo Del Sistema CardiovascularLupita NegronAún no hay calificaciones
- Actividad 1. Embriologia CardiovascularDocumento10 páginasActividad 1. Embriologia Cardiovascularkarime marquezAún no hay calificaciones
- Guía de Práctica 12 2023Documento13 páginasGuía de Práctica 12 2023JAROLL JHANPOLL CUZCO MENDOZAAún no hay calificaciones
- Cap#13 - Sistema CardiovascularDocumento19 páginasCap#13 - Sistema CardiovascularTicci LopezAún no hay calificaciones
- Resumen Sistema CardiovascularDocumento10 páginasResumen Sistema Cardiovascularalexandersamuel Ance GarciaAún no hay calificaciones
- Anatomía Del Desarrollo 2Documento45 páginasAnatomía Del Desarrollo 2nimAún no hay calificaciones
- EMBRIO ResumenDocumento18 páginasEMBRIO Resumencali gonzalezAún no hay calificaciones
- Capitulo 13 - EmbrioDocumento6 páginasCapitulo 13 - EmbrioANA CRISTINA RAMIREZ SIERRAAún no hay calificaciones
- Trabajo de Fisiologia Cardiovascular 5to AñoDocumento20 páginasTrabajo de Fisiologia Cardiovascular 5to AñoMagglis Zerpa AvelloAún no hay calificaciones
- EFAp CV 5 Dr. García 2023-1 PDFDocumento127 páginasEFAp CV 5 Dr. García 2023-1 PDFLuana DAún no hay calificaciones
- Guia Embrio Ordi PalomaDocumento24 páginasGuia Embrio Ordi PalomaDana Paloma GarciaAún no hay calificaciones
- Embriologia de Cardio_100500Documento18 páginasEmbriologia de Cardio_100500AnyelexAún no hay calificaciones
- Informe Caso N°4-Grupo 3Documento9 páginasInforme Caso N°4-Grupo 3SUZANNE FRANCESCA ECHAIZ NARROAún no hay calificaciones
- Embriología Del Sistema CardiovascularDocumento14 páginasEmbriología Del Sistema CardiovascularJazmin AlvarezAún no hay calificaciones
- RESUMEN DEL SISTEMA CARDIOVASCULARDocumento3 páginasRESUMEN DEL SISTEMA CARDIOVASCULARGustavo OjedaAún no hay calificaciones
- Embriología: Sistema Cardiovascular PresentaciónDocumento11 páginasEmbriología: Sistema Cardiovascular PresentaciónEverardo MartinezAún no hay calificaciones
- CorazonDocumento14 páginasCorazonYareli PerezAún no hay calificaciones
- Desarrollo Embriológico Del Corazón y PericardioDocumento6 páginasDesarrollo Embriológico Del Corazón y PericardioYlianaAún no hay calificaciones
- Embriogénesis Del CorazónDocumento5 páginasEmbriogénesis Del CorazónDiana RodriguezAún no hay calificaciones
- Sistema Embriologico CardioRespiratorioDocumento44 páginasSistema Embriologico CardioRespiratorioleonelknightAún no hay calificaciones
- Sistema CardiovascularDocumento11 páginasSistema CardiovascularSol Arteaga RoaAún no hay calificaciones
- Embriología - Sistema CardiovascularDocumento16 páginasEmbriología - Sistema Cardiovascularjohan manuel candelario de la rosaAún no hay calificaciones
- Formación del corazón y circulación fetalDocumento5 páginasFormación del corazón y circulación fetalester saenzAún no hay calificaciones
- Sistema CardiovascularDocumento40 páginasSistema CardiovascularKevin Lopez santa mariaAún no hay calificaciones
- Sistema Cardiocirculatorio YnerviosoDocumento66 páginasSistema Cardiocirculatorio YnerviosoDiego MenaAún no hay calificaciones
- Presentación Libreta Aesthetic Lila PastelDocumento123 páginasPresentación Libreta Aesthetic Lila PastelDani MqzAún no hay calificaciones
- ENSAYODocumento7 páginasENSAYOMayreli Robles De JavierAún no hay calificaciones
- Sistema VascularDocumento6 páginasSistema Vascularmichelle oriana Maqueda TovarAún no hay calificaciones
- Sistema CardiorespiratorioDocumento14 páginasSistema CardiorespiratorioEsteban Ramos MarinAún no hay calificaciones
- GUÍA PRACT 8va SesiónDocumento13 páginasGUÍA PRACT 8va SesiónPablo GGAún no hay calificaciones
- CardioDocumento6 páginasCardiomichelle oriana Maqueda TovarAún no hay calificaciones
- Embriogenesis Del Aparato CardiovascularDocumento15 páginasEmbriogenesis Del Aparato CardiovascularJes inozAún no hay calificaciones
- TALLER Embrio RespuestasDocumento17 páginasTALLER Embrio RespuestasAnge Mendoza AlvarezAún no hay calificaciones
- Desarrollo del sistema cardiovascularDocumento51 páginasDesarrollo del sistema cardiovascularNicole DíazAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento13 páginasEmbriologia Del Sistema CardiovascularMirgelys VargasAún no hay calificaciones
- CAPITULO 13 Embriología LangmanDocumento9 páginasCAPITULO 13 Embriología LangmanLuis Fernando Toro Hernandez100% (6)
- Resumen de Cardiologia-1Documento5 páginasResumen de Cardiologia-1jennifer garciaAún no hay calificaciones
- Resumen Capitulo 17 Langman - Formación Del Sistema CardiacoDocumento5 páginasResumen Capitulo 17 Langman - Formación Del Sistema Cardiacorafael mendezAún no hay calificaciones
- EMBRIOLOGÍA 12 CARDIOVASCULARDocumento87 páginasEMBRIOLOGÍA 12 CARDIOVASCULARvanesa.carranza10Aún no hay calificaciones
- Sistema CardiovascularDocumento20 páginasSistema CardiovascularmaggyemmelyAún no hay calificaciones
- Gutierrez Herrera ResumenDocumento3 páginasGutierrez Herrera ResumenErnesto Antonio Gutierrez HerreraAún no hay calificaciones
- Gutierrez Herrera ResumenDocumento3 páginasGutierrez Herrera ResumenErnesto Antonio Gutierrez HerreraAún no hay calificaciones
- 2.5.2 Tabla Vías MetabólicasDocumento6 páginas2.5.2 Tabla Vías MetabólicasErnesto Antonio Gutierrez HerreraAún no hay calificaciones
- 412-1 Taller de Biocel MEDICINADocumento3 páginas412-1 Taller de Biocel MEDICINAErnesto Antonio Gutierrez HerreraAún no hay calificaciones
- Exploración Por DermatomasDocumento1 páginaExploración Por DermatomasEnriqueRamosAún no hay calificaciones
- Practica Calificada N°1 GeneralidadesDocumento5 páginasPractica Calificada N°1 GeneralidadesTio BeniAún no hay calificaciones
- Cuestionario 8 Sistema Respiratorio Cartaceña Iara-Com.8Documento2 páginasCuestionario 8 Sistema Respiratorio Cartaceña Iara-Com.8iara cartaceñaAún no hay calificaciones
- Fronto PlastiaDocumento57 páginasFronto Plastiagregorio caballeroAún no hay calificaciones
- Sistema osteomuscular funcionesDocumento4 páginasSistema osteomuscular funcionesNicole AnelysAún no hay calificaciones
- Sist. Gen. FemDocumento8 páginasSist. Gen. FemJhoana VazquezAún no hay calificaciones
- Capítulo 001Documento21 páginasCapítulo 001Deceno COLAún no hay calificaciones
- Organos LinfaticosDocumento48 páginasOrganos Linfaticosrubyrespaldo00Aún no hay calificaciones
- Cuestionario Histologia PDFDocumento6 páginasCuestionario Histologia PDFlucasAún no hay calificaciones
- Trabajo de Morfofisiologia AnimalDocumento7 páginasTrabajo de Morfofisiologia Animaljaiver andres claro ojedaAún no hay calificaciones
- Sistema Respitario, Porcion Conductora, Porcion RespiratoriaDocumento55 páginasSistema Respitario, Porcion Conductora, Porcion RespiratoriaCarlos Walter Leon GarridoAún no hay calificaciones
- Infografia CardiovascularDocumento3 páginasInfografia CardiovascularlguillenarciniegaAún no hay calificaciones
- Anatomía TopográficaDocumento6 páginasAnatomía TopográficaMelissa Tello vargasAún no hay calificaciones
- 1º Examen Inglés 1 EvaluaciónDocumento3 páginas1º Examen Inglés 1 EvaluaciónSergio Garcia Isabel BerenguelAún no hay calificaciones
- Anatomia Del AveDocumento24 páginasAnatomia Del AveGeo GutiérrezAún no hay calificaciones
- Tarea CelulasDocumento4 páginasTarea CelulasEsmeralda DávilaAún no hay calificaciones
- A2. 3ra A 8va Semana 2P - GBD 1010Documento6 páginasA2. 3ra A 8va Semana 2P - GBD 1010Guillermo Burgos DiazAún no hay calificaciones
- Grupo N°1 - Tejido EpitelialDocumento2 páginasGrupo N°1 - Tejido EpitelialMaryury RiosAún no hay calificaciones
- Resumen Sistema LinfaticoDocumento13 páginasResumen Sistema LinfaticoMARYLIN GABRIELA PAX TEYULAún no hay calificaciones
- Práctica Del Sistema ÓseoDocumento8 páginasPráctica Del Sistema ÓseoOdaliza Estefani MonesAún no hay calificaciones
- Fisiología cardiovascular: Curso de fisiología humanaDocumento37 páginasFisiología cardiovascular: Curso de fisiología humanaWilder Alex Espinola MariñosAún no hay calificaciones
- Cuaderno de Trabajo BgnRoeoDocumento124 páginasCuaderno de Trabajo BgnRoeoEsdras Suazo SuazoAún no hay calificaciones
- Huesos de La Cara - AlvaroDocumento4 páginasHuesos de La Cara - AlvaroCristian TorrezAún no hay calificaciones
- Sistemas circulatorio, respiratorio y digestivo del delfínDocumento2 páginasSistemas circulatorio, respiratorio y digestivo del delfínAriana Priscilla Velasquez VelascoAún no hay calificaciones
- Los Organos Del Cuerpo Humano 2DO BDocumento9 páginasLos Organos Del Cuerpo Humano 2DO BYenny Marelis LunaAún no hay calificaciones
- Sistema tegumentario, esquelético y muscular del conejo comúnDocumento16 páginasSistema tegumentario, esquelético y muscular del conejo comúnNaleibis NievesAún no hay calificaciones
- ESTOMAGO - Exploracion FisicaDocumento13 páginasESTOMAGO - Exploracion FisicaCarmen TorresAún no hay calificaciones
- Semana 8 1Documento27 páginasSemana 8 1ANGELA GUERRERO MARTINEZAún no hay calificaciones
- Josue David Díaz Gutiérrez - PRACTICA-20.-TEJIDO-OSEODocumento6 páginasJosue David Díaz Gutiérrez - PRACTICA-20.-TEJIDO-OSEOMARIANA VALENTINA GARCIA GONZALEZAún no hay calificaciones
- Guía 13 DigestivoDocumento3 páginasGuía 13 Digestivolouis tomlinsonAún no hay calificaciones