Documentos de Académico
Documentos de Profesional
Documentos de Cultura
Sistema Endocrino
Cargado por
Amadeo CabelloTítulo original
Derechos de autor
Formatos disponibles
Compartir este documento
Compartir o incrustar documentos
¿Le pareció útil este documento?
¿Este contenido es inapropiado?
Denunciar este documentoCopyright:
Formatos disponibles
Sistema Endocrino
Cargado por
Amadeo CabelloCopyright:
Formatos disponibles
SISTEMA ENDOCRINO
1.-INTRODUCCIÓN
Es uno de los sistemas de regulación y control del cuerpo humano. Su función principal es coordinar
y regular diversas funciones del organismo a través de la secreción de sustancias químicas llamadas
hormonas. Estas hormonas son producidas por glándulas endocrinas y son liberadas en el torrente sanguíneo,
desde donde pueden afectar a células y tejidos en todo el cuerpo. El sistema endocrino es una colección de
glándulas cuya función es regular múltiples órganos dentro del organismo para: a) satisfacer las necesidades
de crecimiento y reproducción del organismo, y b) responder a las fluctuaciones del ambiente interno,
incluidos diversos tipos de estrés. El sistema endocrino incluye las siguientes glándulas principales:
• Páncreas endocrino.
• Glándulas paratiroides.
• Glándula pituitaria (hipófisis, que se asocia con los núcleos
hipotalámicos).
• Tiroides.
• Glándulas suprarrenales.
• Gónadas (testículos y ovarios)
Estas glándulas endocrinas sintetizan y secretan hormonas bioactivas y,
con excepción de las gónadas, que realizan funciones tanto endocrinas como gametogénicas, se dedican en
exclusiva a la producción hormonal. Un órgano transitorio, la placenta, también realiza una importante
función endocrina.
Además de las glándulas endocrinas exclusivas, existen células endocrinas en el seno de otros órganos, cuya
función principal no es endocrina. Entre ellas destacan las células del corazón productoras de péptido
natriurético atrial, las células hepáticas productoras del factor del crecimiento parecido a la insulina I (IGF-
I), las células renales productoras de eritropoyetina, y numerosos tipos celulares en el tubo digestivo que
producen hormonas digestivas..
Un tercer brazo del sistema endocrino está representado por numerosos tipos celulares que expresan enzimas
intracelulares, ect. enzimas secretadas, que modifican precursores inactivos u hormonas menos activas para
dar lugar a hormonas muy activas. Un ejemplo puede ser la formación de angiotensina-II.
La transmisión de señales endocrinas implica: a) la secreción regulada de una molécula transmisora de
señales extracelulares, denominada hormona, al líquido extracelular; b) la difusión de la hormona hacia los
vasos y su circulación por todo el organismo, y c) la difusión de la hormona fuera del compartimento
vascular hacia el espacio extracelular, con unión a un receptor específico dentro de las células del órgano
diana.
2.- EJE HIPOTÁLAMO-HIPÓFISIS
Hipotálamo Adenohipófisis Glándula diana Efectos biológicos
Hormona Hormona Hormona de la
liberador trópica glándula diana
a
3.-EJE HIPOTÁLAMO-HIPÓFISIS-TIROIDEO
3.1.- Glándula tiroides
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
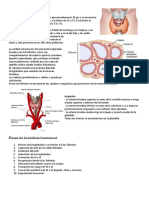
Situada en el cuello, por debajo del cartílago cricoides, constituida
por dos lóbulos unidos por el istmo del tiroides. Su unidad
funcional es el folículo tiroideo el cual está constituido por:
• Células foliculares: encargadas de la síntesis de hormonas
tiroideas
• Células C (parafoliculares): sintetizan calcitonina
• Coloide: sustancia gelatinosa que llena la cavidad central del
folículo tiroideo. Está compuesto principalmente de tiroglobulina,
que es una proteína especializada en la síntesis y almacenamiento
de las hormonas tiroideas.
3.2.- Hormonas tiroideas.
• T3/Triyodotironina: tiene mayor actividad que la T4
• T4/ Tiroxina: menor actividad que la anterior, al llegar a la célula diana se convierte en T3.
3.3.- Síntesis de hormonas tiroideas.
1. Captación de I- (transporte secundario con Na+). Esto se produce gracias a que la TSH facilita su
entrada a través de NIS
2. Transporte de yodo al coloide a través de pendrina
3. Síntesis de tiroglobulina
4. Oxidación del yodo por peroxidasas
5. Iodación de Tiroxina: formación de T1 y T2
6. Acoplamiento de Tiroxina: formación de
a. T3 (T2+T1) Triyodotironina
b. T4 (T2+T2): Tiroxina
7. Almacenamiento de hormonas acopladas a Tiroglobulina
8. Por pinocitosis salen del coloides hacia las células foliculares
9. Degradación de TG+T1+T2+T3+T4 por proteasas (eliminando tiroglobulina)
10. Desyodación. Reciclaje yodo por la desyodasa
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
3.4.-Liberación de hormonas tiroideas
1. El proceso de señalización que conduce a la liberación de T3 (triiodotironina) y T4 (tiroxina)
comienza con la estimulación del hipotálamo, que secreta la hormona liberadora de tirotropina
(TRH).
2. Luego, la adenohipófisis (parte anterior de la glándula pituitaria) produce la hormona estimulante de
la tiroides (TSH) en respuesta a la TRH.
3. La TSH viaja a través del torrente sanguíneo y llega a la glándula tiroides. En la superficie de las
células foliculares en la tiroides, la TSH se une a receptores específicos acoplados a proteína G. La
unión a este receptor da lugar a:
a. Activación de PLC que rompe PIP2 en DAG y en IP3 que actúa sobre receptores
sarcoplásmicos produciendo la liberación de iones calcio que potencian la liberación de TSH.
b. Activación de la AC que convierte el ATP (adenosín trifosfato) en AMPc cuyo aumento
potencia la estimulación de la síntesis y liberación de las hormonas tiroideas T3 y T4.
4. Síntesis y liberación de T3 y T4 al torrente sanguíneo.
3.5.- Transporte de hormonas tiroideas.
La mayoría de las hormonas tiroideas liberadas en el torrente sanguíneo se unen a proteínas transportadoras
específicas. Las dos principales proteínas transportadoras son la globulina fijadora de tiroxina (TBG) y la
transtiretina (TTR), también llamada prealbúmina. Estas proteínas se producen en el hígado y transportan las
hormonas tiroideas en el torrente sanguíneo. También se unen a la albúmina.
Las hormonas tiroideas, unidas a las proteínas transportadoras, circulan por todo el cuerpo. La unión a estas
proteínas transportadoras aumenta la solubilidad de las hormonas tiroideas en el agua y prolonga su vida
media en la sangre.
3.6.- Eliminación de hormonas tiroideas.
La eliminación de las hormonas tiroideas T3 (triiodotironina) y T4 (tiroxina) del organismo está regulada
principalmente por un grupo de enzimas llamadas desyodasas. Hay tres tipos principales de desyodasas: D1,
D2 y D3, y cada una tiene un papel específico en la regulación de las hormonas tiroideas.
• Desyodasa Tipo 1 (D1):
o Ubicación: en los tejidos de alto flujo con intercambios rápidos con el plasma, como el
hígado, los riñones y el músculo esquelético
o Función: La D1 tiene un papel fundamental en la conversión de la hormona tiroidea T4 en la
forma más activa, T3. Esto se conoce como la conversión periférica de T4 a T3.
• Desyodasa Tipo 2 (D2):
o Ubicación: La D2 se encuentra principalmente en tejidos como SNC, células tirotropas de la
hipófisis y placenta.
o Función: La D2 es especialmente importante en la conversión local de T4 a T3 dentro de
estos tejidos. Esto permite un control más específico de los niveles de T3 en los tejidos donde
es necesario para la regulación de funciones específicas.
• Desyodasa Tipo 3 (D3): convierte T4 en rT3 inactiva. La desyodinasa de tipo 3 aumenta durante el
hipertiroidismo, lo que ayuda a amortiguar la sobreproducción de T4. Todas las formas de
yodotironinas se desyodan más, hasta producir tironinas no yodadas
3.7.- Mecanismo de acción de hormonas tiroideas.
Ejercen sus efectos a través de mecanismos de acción tanto genómicos como no genómicos en las células
objetivo. Estos mecanismos permiten a las hormonas tiroideas regular una variedad de funciones fisiológicas
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
y metabólicas en el cuerpo.
A. Mecanismo de Acción Genómica
a. T4 y T3 entran en la célula por mediación de transportadores o por difusión.
b. En los tejidos diana la 5´/3´monodesyodasa transforma T4 en T3
c. El receptor de dichas moléculas no se encuentra en el citoplasma sino que se encuentra en el
núcleo.
d. Estas hormonas al ser liposolubles atraviesan las membranas con facilidad (plasmática y
nuclear) y se une con su receptor, receptor de hormonas tiroideas (TR), heterodímero el cual
está unido al elemento de respuesta tiroidea (TRE) y a un receptor de ácido retinoico (RXR)
Los TR existen en dos isoformas principales: TR alfa y TR beta
e. Tras la unión, el TRE se activa y permite que la caja TATA se estabilice y se lleva a cabo la
transcripción dando lugar a ARNm que posteriormente permitirá la formación de proteínas.
INDUCCIÓN DE GENES
f. Efectos Metabólicos y Fisiológicos: Las proteínas producidas en respuesta a la activación del
gen diana tienen un impacto significativo en el metabolismo y las funciones celulares.
g. También puede existir el elemento de respuesta tiroidea negativo que no permite que se forme
el ARNm y, en consecuencia, no se forman proteínas. (no es muy común en este tipo de
hormonas)
B. Mecanismo de Acción No Genómica:
Además de los efectos genómicos, las hormonas tiroideas también pueden tener efectos no genómicos que
ocurren rápidamente y no implican la regulación de genes
a. Acción en la Membrana Celular: Las hormonas tiroideas pueden influir en la membrana
celular, alterando la permeabilidad y la excitabilidad de las células. Esto puede afectar la
conducción de señales nerviosas y la contracción muscular.
b. Regulación de Canales Iónicos: Las hormonas tiroideas pueden modular la actividad de
canales iónicos en la membrana celular. Esto puede tener efectos significativos en la
excitabilidad de las células y la transmisión de señales.
c. Interacción con Proteínas de Membrana: Además, las hormonas tiroideas pueden interactuar
directamente con proteínas de membrana específicas, como transportadores de nutrientes y
receptores de superficie. Esto puede influir en la absorción de nutrientes y la regulación de la
homeostasis de iones
d. Respuesta Rápida en Tejidos Específicos: La respuesta rápida de las hormonas tiroideas es
particularmente importante en tejidos que requieren adaptaciones rápidas, como el corazón y
el músculo esquelético. En el corazón, por ejemplo, la activación rápida de canales iónicos y
vías de señalización por parte de las hormonas tiroideas puede aumentar la contractilidad
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
cardíaca y la frecuencia cardíaca en cuestión de segundos.
i. Aumento del metabolismo basal:
1. Aumento de la Lipólisis: Las hormonas tiroideas estimulan la lipólisis, que es
el proceso de descomposición de las grasas almacenadas en el tejido adiposo
(grasa corporal) en ácidos grasos y glicerol que pueden ser utilizados como
energía o sustratos para llevar a cabo la gluconeogénesis
2. Aumentan la proteólisis. Este aumento en la proteólisis se debe a la capacidad
de las hormonas tiroideas de aumentar la tasa metabólica general del
organismo.
Cuando las hormonas tiroideas están en niveles elevados, aumenta la tasa metabólica basal, lo que significa
que el cuerpo quema más energía (calorías) incluso en reposo. Para mantener esta mayor tasa metabólica, el
cuerpo puede movilizar una variedad de fuentes de energía, incluyendo las grasas y las proteínas. En el caso
de las proteínas, esto implica la proteólisis, donde se descomponen las proteínas musculares y otras
proteínas en el cuerpo para obtener aminoácidos que pueden ser utilizados como fuente de energía. Si bien
este aumento en la proteólisis puede contribuir a la pérdida de peso en personas con hipertiroidismo (niveles
elevados de hormonas tiroideas), también puede llevar a la pérdida de masa muscular y a una disminución
de la fuerza. Esto puede ser perjudicial para la salud a largo plazo.
3. Aumento del funcionamiento de la bomba de sodio-potasio (Na+/K+ ATPasa)
en las células.
La bomba trabaja de manera más eficiente para bombear sodio fuera de la célula y potasio dentro de la
célula. El aumento en la actividad de la bomba de sodio-potasio requiere energía en forma de ATP (adenosín
trifosfato). Como resultado, el aumento de la actividad de esta bomba puede contribuir al aumento general
en el metabolismo y el gasto de energía observado en personas con niveles elevados de hormonas tiroideas
(hipertiroidismo).
3.8.- Efectos fisiológicos de las hormonas tiroideas.
• Aumento del Metabolismo Basal: Las hormonas tiroideas estimulan la actividad de la citocromo c
oxidasa y la deshidrogenasa en las mitocondrias, lo que potencia la oxidación de glucosa y ácidos
grasos, incrementando la producción de trifosfato de adenosina (ATP) y el consumo de oxígeno,
resultando en un aumento del metabolismo basal.
• Efectos Cardiovasculares: Promueven la expresión de genes que codifican proteínas contractiles
cardíacas, como la miosina, mejorando la contractilidad miocárdica. También incrementan la
sensibilidad de los receptores adrenérgicos beta y alfa, lo que aumenta la respuesta a catecolaminas y
contribuye al gasto cardíaco y la perfusión tisular.
• Estimulación del Sistema Nervioso Central: A nivel del sistema nervioso central, las hormonas
tiroideas intensifican la síntesis y liberación de neurotransmisores, como la serotonina y
noradrenalina, modulando la excitabilidad neuronal, lo que potencia la concentración y el estado de
alerta.
• Metabolismo de Carbohidratos y Lípidos: Incrementan la expresión de transportadores de glucosa y
enzimas implicadas en la glucólisis y la oxidación de ácidos grasos, estimulando la captación de
glucosa y la oxidación de lípidos en las mitocondrias, regulando así los niveles de glucosa en sangre
y la lipólisis.
• Aumento de la Ventilación Pulmonar: Mediante un aumento de la demanda de oxígeno tisular, las
hormonas tiroideas estimulan la ventilación pulmonar, generando un aumento de la frecuencia y
profundidad respiratoria, lo que asegura un aporte adecuado de oxígeno.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• Crecimiento y Desarrollo: Son esenciales para el desarrollo normal del sistema nervioso central y
regulan el cierre de las placas epifisarias en los huesos largos, lo que influye en el crecimiento y la
estructura ósea.
• Termorregulación: Estimulan la termogénesis y aumentan la sensibilidad de los receptores de
temperatura, fomentando la producción de calor en tejidos periféricos y la capacidad de respuesta al
frío.
• Modulación Inmunológica: Pueden influir en la proliferación linfocitaria y la producción de
citocinas, ejerciendo un efecto en la respuesta inmunológica y la inflamación.
• Efectos en el Tracto Gastrointestinal: Incrementan la motilidad gastrointestinal y pueden influenciar
la absorción de nutrientes y la función intestinal, impactando la digestión y el metabolismo de
nutrientes.
4.- ALTERACIONES DE LA ACTIVIDAD TIROIDEA
4.1.-Bocio
Dentro de las alteraciones morfológicas encontramos el bocio que es un aumento de la propia glándula,
puede estar causada por:
• Un problema de falta de yodo donde el tiroides aumenta el tamaño para equilibrar el déficit.
• Hay otras veces que puede aumentar porque nos encontramos un problema en la secreción de
tiroxina.
Un bocio simple ocurre por una baja ingesta de yodo o alteración que disminuye su actividad, cuando
hablamos de bocio multinodular es que aparecen nódulos o tumoración y esta parte aumenta el tamaño.
Bocio difuso o simple es cuando hay un aumento general y proporcional, relacionado con deficiencia de
yodo o por poemas autoinmunes. Cuando la alteración tiroidea se refiere a un nódulo entonces se denomina
nodular porque hay un aumento de la zona donde se desarrolla el nódulo. Hay veces que el bocio no se
desarrolla por hiper o hipotiroidismo.
4.2.-Hipotiroidismo.
Insuficiencia de la hormona tiroidea, que origina un enlentecimiento general de la tasa metabólica. Hay
sitios donde el hipotiroidismo también está muy relacionado con la dieta, porque si no se toma alimentos con
yodo la probabilidad de sufrir la enfermedad aumenta.
Las causas son:
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• Primarias: defecto en la propia glándula:
o Atrofia congénita
o Deficiencia de yodo o Tiroiditis de Hashimoto o enfermedad autoinmune, consecuencia de la
enfermedad de Graves
• Secundarias
o Defecto hipofisario
o Terciarias:
▪ Defecto hipotalámico
▪ Yatrogénicas
▪ Tratamiento con yodo
▪ Cirugía del tiroides
▪ Drogas antitiroideas
Tratamiento: L-tiroxina
4.3.- Cretinismo
Trastorno congénito poco común que se produce debido a la deficiencia grave de hormonas tiroideas en el
período prenatal o durante la infancia temprana. Las hormonas tiroideas son esenciales para el desarrollo
adecuado del cerebro y el crecimiento del cuerpo en los primeros años de vida. Cuando hay una deficiencia
grave de estas hormonas en esta etapa crucial, puede dar lugar al cretinismo. Algunas de las manifestaciones
más comunes incluyen (no importante):
• Retraso en el Desarrollo Físico y Mental: Esto puede incluir un crecimiento lento, retraso en la
adquisición de habilidades motoras y un desarrollo cognitivo limitado.
• Retraso en el Desarrollo Óseo: Los huesos pueden ser más cortos y menos densos de lo normal.
• Rostro Peculiar: Características faciales típicas del cretinismo incluyen un puente nasal bajo, una
nariz achatada, labios gruesos, párpados hinchados y una lengua grande.
• Retardo Mental: La deficiencia de hormonas tiroideas en el período prenatal o durante la infancia
temprana puede llevar a retraso mental y dificultades de aprendizaje.
• Dificultades de Habla y Audición: Pueden presentarse problemas en el habla y la audición.
• Problemas en la Piel y el Cabello: La piel puede ser seca y escamosa, y el cabello puede ser fino y
quebradizo.
El tratamiento con hormona tiroidea da lugar al “Crecimiento sustitivo” Sin embargo el tratamiento solo
mejora el desarrollo mental en caso de que se instaure en los primeros días de vida.
4.4.- Hipertiroidismo
Trastorno de la glándula tiroides en el cual esta produce y libera en exceso hormonas tiroideas. Los
principales síntomas son: (no importante)
• Pérdida de peso no intencionada.
• Aumento de la frecuencia cardíaca (taquicardia) y palpitaciones.
• Nerviosismo y ansiedad.
• Temblores en las manos.
• Sudoración excesiva y sensibilidad al calor.
• Fatiga y debilidad muscular.
• Cambios en la piel, como adelgazamiento y debilitamiento del cabello.
• Problemas para dormir (insomnio).
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• Cambios en el ciclo menstrual en mujeres.
• Diarrea o movimientos intestinales frecuentes.
• Hinchazón en el cuello debido al agrandamiento de la tiroides (bocio)
Las causas principales son:
• Hiperactividad del tiroides, ha aumentado la prevalencia en los últimos años sobre todo por causas
autoinmunes que provoca que se creen unos anticuerpos que activan los órganos diana para que se
produzca T3 y T4, este proceso recibe el nombre específico de enfermedad de graves.
• Respuesta acelerada del tiroides por un proceso infeccioso, denomina tiroiditis. Las causas
secundarias ocurren por problemas de estimulación en personas que tengan un tumor hipofisario.
• Tratamiento farmacológico, que tenga como principio activo el yodo (como por ejemplo litio) que
sería un tipo de causa yatrogénica.
Se puede tratar con:
• Inhibidores de Receptores β1 Adrenérgicos: Estos medicamentos, como el propranolol, bloquean los
receptores β1 adrenérgicos en el corazón y se utilizan principalmente para controlar los síntomas de
la hiperactividad del sistema nervioso simpático, que a menudo se observa en el hipertiroidismo..
• Fármacos Antitiroideos: Los fármacos antitiroideos, como el metimazol o el propiltiouracilo, se
utilizan para tratar el hipertiroidismo. Estos medicamentos reducen la producción de hormonas
tiroideas en la glándula tiroides y ayudan a controlar el exceso de actividad tiroidea. El objetivo del
tratamiento es normalizar la función tiroidea y restaurar el eutiroidismo.
• Yodo Radiactivo: El yodo radioactivo se utiliza para tratar el hipertiroidismo, especialmente en casos
de enfermedad de Graves. El yodo radioactivo se acumula en la glándula tiroides y destruye
selectivamente las células tiroideas hiperactivas. El objetivo es reducir la actividad tiroidea excesiva
y lograr la eutiroidismo.
4.5.- Enfermedad de Graves.
También conocida como enfermedad de Basedow-Graves o tirotoxicosis autoinmunitaria, es una enfermedad
autoinmunitaria que afecta la glándula tiroides. En esta afección, el sistema inmunológico del cuerpo ataca
la glándula tiroides y provoca una sobreproducción de hormonas tiroideas. Se caracteriza por la producción
de TSI, anticuerpos estimulantes del receptor de la tirotropina (TRAb), o anticuerpos antirreceptor de la
tirotropina (TSH) en el cuerpo. Estos anticuerpos imitan la acción de la hormona estimulante de la tiroides
(TSH), que normalmente regula la producción de hormonas tiroideas en la glándula tiroides. Sin embargo,
en la enfermedad de Graves, los anticuerpos activan en exceso los receptores de TSH en la tiroides, lo que
lleva a una producción excesiva de hormonas tiroideas.
4.6.- Efectos de la hormona tiroidea en la expresión génica cardíaca
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
4.7.- Tiroticoxis y corazón
Se entiende por tirotoxicosis al exceso de HT circulantes, sean estas de origen exógeno (p. ej., dosis
exageradamente altas de HT prescritas inadvertidamente por el medico) o bien de procedencia endógena. En
este último caso se habla con propiedad de hipertiroidismo. Los pacientes con tirotoxicosis se presentan,
clínicamente, con síntomas y signos característicos, muchos de ellos referidos al corazón y al sistema
cardiovascular.
4.8.- Hipotiroidismo y corazón.
Los síntomas y signos cardiovasculares del hipotiroidismo son, en general, opuestos a los del
hipertiroidismo
Cuando se presenta una disminución en la función tiroidea, se pueden observar varios hallazgos médicos.
Entre ellos, se encuentra la bradicardia sinusal, que se caracteriza por una frecuencia cardíaca más lenta de
lo normal. También puede notarse una leve hipertensión arterial con un predominio en la fase diastólica del
ciclo cardíaco, lo que se traduce en una reducción en la diferencia entre la presión arterial sistólica y
diastólica, debido a un aumento en la resistencia de los vasos sanguíneos en el sistema circulatorio.
Además, los pacientes pueden experimentar fatiga y sensibilidad al frío. A nivel cardíaco, se observa una
disminución en la capacidad de contracción del corazón y, como consecuencia, una disminución en el
volumen de sangre bombeado por minuto, lo que se conoce como gasto cardíaco reducido. Esta disminución
en la capacidad de contracción se debe, en parte, a cambios en la expresión genética causados por la
deficiencia de hormonas tiroideas (HT). Dichos cambios afectan negativamente a la actividad de una enzima
llamada Ca2+ - ATPasa del retículo sarcoplásmico, así como a un incremento en un inhibidor de esta enzima
denominado fosfolamban. Ambas proteínas son fundamentales en la regulación de los niveles de calcio
dentro de las células cardíacas, y el calcio es esencial para el proceso de contracción y relajación del
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
músculo cardíaco, influyendo tanto en la función contráctil durante la eyección de sangre (sístole) como en
la relajación durante el llenado de las cavidades cardíacas (diástole).
Adicionalmente, el hipotiroidismo provoca alteraciones en el metabolismo de las lipoproteínas en la sangre,
lo que conduce a niveles elevados de colesterol en la forma de lipoproteína de baja densidad (LDL) y
apolipoproteína B. Este proceso contribuye a la aceleración de la aterosclerosis, que es el endurecimiento y
estrechamiento de las arterias debido a la acumulación de placa. La base subyacente de este fenómeno se
relaciona con la disminución tanto en la cantidad como en la actividad de los receptores hepáticos para el
LDL, lo que resulta en una reducción en la eliminación del colesterol LDL de la sangre
Claves para el examen (Clase):
- ¿Cómo llega la TRH a la adenohipófisis? sistema porta-hipofisario
- ¿Qué estímulo necesita la adenohipófisis para secretar TSH a la circulación?
Incremento de calcio (IP3, PLC, PG).
- ¿Qué, quién, cuando y por qué?
- ¿Qué necesita la tiroides para producir tiroxina? Yodo, NIS, Pendrina.
5.- GLÁNDULAS PARATIROIDEAS.
Los seres humanos poseen cuatro glándulas paratiroides, dos situadas en la superficie posterior del lóbulo
izquierdo de la tiroides y otras dos en el derecho. En conjunto, estas cuatro glándulas pesan menos de
500 mg. Están compuestas en su mayor parte por células principales, que son las responsables de sintetizar
y secretar PTH. Estas células, como las que secretan otras hormonas peptídicas, están altamente
especializadas en sintetizar, procesar y secretar su producto. El principal factor regulador de la secreción de
PTH es el Ca2+ iónico plasmático, aunque la vitamina D también participa. Ambos inhiben la síntesis o
liberación de PTH. Por el contrario, un aumento en la concentración plasmática de fósforo estimula la
liberación de PTH
5.1.- Síntesis de PTH.
Se lleva a cabo en las células principales:
1. En primer lugar se produce la transcripción del Gen
2. Seguido de la síntesis del ARN Mensajero (ARNm)
con la posterior traducción del ARNm.
3. Tras esto, se produce el procesamiento
preprohormona PTH; la PTH se sintetiza
inicialmente como una prohormona más grande
llamada "preprohormona". Esta preprohormona
contiene una secuencia de señal que dirige la
molécula hacia el retículo endoplasmático rugoso
(RER), donde se produce el procesamiento inicial.
En el RER, la preprohormona experimenta varios
procesos importantes:
o Eliminación de la Secuencia de Señal: La preprohormona contiene una secuencia de señal en
su extremo N-terminal que es necesaria para dirigir la molécula al RER. En este punto, se
elimina esta secuencia de señal, lo que da como resultado una molécula más corta llamada
"prohormona".
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
o Modificaciones Post-Translacionales: Durante su paso por el RER, la prohormona puede
someterse a modificaciones post-traduccionales, como la adición de grupos glucosa o la
formación de enlaces disulfuro, que son necesarias para su correcto plegamiento y
procesamiento.
4. Empaquetamiento en Vesículas Secretoras y Transporte al Aparato de Golgi: La prohormona es
empaquetada en vesículas secretoras y transportada al aparato de Golgi, donde se somete a procesos
de maduración y procesamiento adicionales.
5. Procesamiento en el Aparato de Golgi: En el aparato de Golgi, la prohormona se modifica y procesa
aún más. Esto incluye la división de la prohormona en fragmentos más pequeños, uno de los cuales
es la hormona activa PTH.
6. Secreción de PTH: Finalmente, las vesículas secretoras que contienen la PTH y otros productos de
procesamiento se fusionan con la membrana celular de las células paratiroideas, permitiendo la
liberación de la PTH madura y activa al torrente sanguíneo.
5.2.- Control de la síntesis de PTH por calcio
La inhibición de la síntesis y liberación de la hormona paratiroidea (PTH) en respuesta a niveles altos de
calcio en sangre se produce a través de un proceso de retroalimentación negativa que involucra
principalmente los receptores de calcio-sensing (CaSR) en la membrana de las células principales de las
glándulas paratiroidea los cuales están acoplados a proteínas G. Descripción más detallada de este proceso:
1. Unión de Calcio a los CaSR: Cuando los niveles de calcio en el entorno extracelular aumentan, las
moléculas de calcio se unen a los sitios específicos en los CaSR.
2. Activación de Proteínas G
3. Activación de la Fosfolipasa C (PLC):
a. La subunidad alfa de la proteína G activada (Gα) interactúa con la fosfolipasa C (PLC)
4. Generación de IP3 y DAG:
a. La PLC divide el PIP2 en IP3 y DAG.
b. El IP3 se libera en el citoplasma y actúa como un
segundo mensajero al unirse a receptores en el
retículo endoplasmático, lo que provoca la liberación
de calcio almacenado en las reservas intracelulares.
c. El DAG permanece en la membrana celular y activa
la proteína quinasa C (PKC)
5. Modulación de la Actividad de Factores de Transcripción:
a. Los niveles elevados de calcio intracelular y la
activación de PKC pueden afectar a factores de
transcripción y reguladores que controlan la síntesis
de PTH.
6. Inhibición de la Transcripción del Gen de la PTH:
a. La fosforilación inducida por PKC y otros eventos intracelulares resulta en la inhibición de la
actividad de estos factores de transcripción.
b. Como resultado, la transcripción del gen de la PTH se reduce significativamente, lo que
significa que se produce menos ARNm de PTH.
7. Disminución de la Síntesis de PTH:
a. Con una reducción en la transcripción del gen de la PTH, se sintetiza menos PTH en las
células principales de las glándulas paratiroideas.
b. La PTH sintetizada se almacena en vesículas secretoras y se libera en menor cantidad al
torrente sanguíneo.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
5.3.- Control de la síntesis de PTH por vitamina D
El gen de la hormona paratiroidea (PTH) está regulado a nivel
transcripcional por los elementos de respuesta a las vitaminas D y A,
específicamente el elemento de respuesta a la vitamina D y su interacción
con el receptor de vitamina D (VDR):
1. Elemento de Respuesta a la Vitamina D en el Gen de la PTH: El gen
de la hormona paratiroidea (PTH) contiene elementos reguladores en su
región upstream o corriente arriba, es decir, antes del inicio de la secuencia
de ADN que codifica la PTH. Entre estos elementos reguladores se
encuentra uno que responde a la vitamina D, conocido como elemento de
respuesta a la vitamina D.
2. Receptor de Vitamina D (VDR): El VDR es un receptor nuclear que pertenece a la familia de
receptores nucleares, similar a los receptores de hormonas esteroides y tiroideas. El VDR tiene la
capacidad de unirse a un metabolito activo de la vitamina D, generalmente el 1,25-dihidroxivitamina
D, que se forma en el organismo.
3. Formación de un Complejo VDR-Vitamina D: Cuando el VDR se une al 1,25-dihidroxivitamina D,
forma un complejo VDR-vitamina D.
4. Heterodimerización con el Receptor X de Retinoides (RXR): El VDR, al igual que el receptor de
hormona tiroidea, forma un heterodímero con el receptor X de retinoides (RXR). Esto significa que
ambos receptores se unen y trabajan juntos como una unidad funcional.
5. Acción como Factor de Transcripción: El complejo formado por el VDR y el RXR actúa como un
factor de transcripción. Esto significa que este complejo tiene la capacidad de influir en la
transcripción de genes específicos.
6. Reducción de la Velocidad de Transcripción de la PTH: Cuando el complejo VDR-vitamina D se une
al elemento de respuesta a la vitamina D en el gen de la PTH, se produce una modulación de la
actividad transcripcional. En este contexto, la unión de este complejo reduce la velocidad de
transcripción del gen de la PTH.
7. Regulación Negativa de la Síntesis de PTH: Esta reducción en la velocidad de transcripción del gen
de la PTH tiene como resultado una disminución en la síntesis de la hormona paratiroidea (PTH).
5.4.- Metabolismo PTH.
Una vez que la PTH es secretada por las células principales de las glándulas paratiroideas, sufre un rápido
proceso de metabolización. Aquí se detallan los aspectos clave de este proceso:
• Vida Media de la PTH: La PTH circula en el plasma sanguíneo y tiene una vida media muy corta de
aproximadamente 4 minutos. Esto significa que se degrada rápidamente en el organismo.
• Escisión en Dos Fragmentos: La PTH se divide en dos fragmentos principales:
o Fragmento N-Terminal: Este fragmento contiene entre 33 y 36 aminoácidos y es esencial para
la actividad biológica de la PTH. Su función principal es regular los niveles de calcio y
fósforo en el cuerpo.
o Fragmento C-Terminal: Este fragmento es más grande y se considera biológicamente
inactivo. A pesar de su falta de actividad, tiene una vida media más larga que el fragmento N-
terminal.
• Inicio de Escisión: La escisión de la PTH comienza en los gránulos secretorios dentro de las células
principales de las glándulas paratiroideas. Posteriormente, este proceso continúa en la circulación
sanguínea, sobre todo en el hígado.
• Hidrólisis en el Riñón: La mayor parte de la actividad biológica de la PTH se encuentra en el
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
fragmento N-terminal. Sin embargo, este fragmento es rápidamente hidrolizado, principalmente en el
riñón. Esto es parte del mecanismo que regula la homeostasis del calcio y el fósforo.
• Vida Media Prolongada del Fragmento C-Terminal: A diferencia del fragmento N-terminal, el
fragmento C-terminal tiene una vida media mucho más larga. Se estima que entre el 70% y el 80%
de la PTH circulante consiste en este fragmento C-terminal, que carece de actividad biológica.
• Dificultades en la Medición: La presencia de una cantidad significativa de fragmentos C-terminales
en la circulación, aunque carecen de actividad biológica, ha complicado la medición precisa de los
niveles de PTH. Esto se debe a que las pruebas tradicionales de radioinmunoensayo pueden detectar
tanto la PTH intacta como los fragmentos C-terminales.
• Técnicas de Medición Específicas: Para abordar esta complicación, se han desarrollado técnicas de
enzimoinmunoensayo (ELISA) que utilizan dos anticuerpos diferentes que se dirigen a puntos
específicos de la molécula de PTH. Esto permite medir solo la cantidad de PTH intacta de 84
aminoácidos, lo que resulta en una medición más precisa de la hormona PTH activa.
5.5.- Efectos de la PTH.
1. Secreción de PTH: las células principales de las glándulas paratiroideas liberan la hormona
paratiroidea (PTH) en la circulación sanguínea.
2. Acción en los Huesos (Efecto Principal): La PTH actúa sobre los huesos de varias maneras:
a. Estimula la formación y activación de osteoclastos, células encargadas de la resorción ósea.
b. Los osteoclastos descomponen la matriz ósea liberando calcio y fósforo en el torrente
sanguíneo, aumentando así los niveles de calcio y fósforo plasmático.
3. Acción en el Riñón:
a. Estimula la conversión de la 25-hidroxivitamina D (forma inactiva de la vitamina D) en su
forma activa, el calcitriol, en los túbulos renales.
b. El calcitriol mejora la absorción intestinal de calcio y fósforo.
c. Aumenta la reabsorción de calcio en los túbulos renales, reduciendo la excreción de calcio en
la orina.
4. Efectos en el Fósforo: Aunque su efecto principal es en el calcio, la PTH también regula los niveles
de fósforo al inhibir su reabsorción en los riñones, lo que lleva a una disminución de los niveles de
fósforo en sangre y a un aumento de los niveles en orina (fosfaturia)
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Así pues, la PTH mantenida desempeña un papel crucial en la regulación del metabolismo óseo,
promoviendo la formación y activación de osteoclastos, células responsables de la resorción ósea. Este
proceso implica la acción de la PTH sobre los osteoblastos y la liberación de una proteína llamada RANKL
(receptor activador del factor nuclear kappa-B ligando) que juega un papel fundamental en la formación de
osteoclastos.
1. Estimulación de Osteoblastos por la PTH: La PTH activa los osteoblastos a través de su receptor
PTH-1R. Como resultado, los osteoblastos liberan moléculas señaladoras, incluida la interleucina 6
(IL-6) y el factor estimulante de colonias de macrófagos (M-CSF), que promueven la formación de
osteoclastos.
2. Producción de RANKL: Los osteoblastos bajo la influencia de la PTH también producen RANKL,
que es una molécula esencial para la formación de osteoclastos. RANKL se une a su receptor RANK
en la superficie de los precursores de osteoclastos, estimulando su diferenciación y activación.
3. Formación de Osteoclastos: Cuando RANKL se une a su receptor RANK en los precursores de
osteoclastos, inicia una cascada de señalización que culmina en la formación y activación de
osteoclastos maduros. Estos osteoclastos están diseñados para resorber la matriz ósea, liberando
calcio y fosfato en la circulación sanguínea.
Este proceso de resorción ósea es fundamental para
mantener el equilibrio de calcio en el organismo y
adaptar la masa ósea a las necesidades cambiantes.
Por otra parte el aumento de los niveles de PTH de
manera intermitente da lugar a síntesis de hueso,
induciendo una mayor velocidad de formación ósea y la
aposición de mineral. El proceso por el cual se produce
es el siguiente:
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
1. Estimulación de los Osteocitos: La PTH
intermitente actúa directamente sobre los osteocitos,
células óseas altamente especializadas ubicadas en las
cavidades óseas. Los osteocitos son clave en la
comunicación y el mantenimiento del tejido óseo.
2. Activación de Canales de Calcio (Ca2+):
La PTH se une a los receptores de PTH en la
membrana de los osteocitos, desencadenando una
cascada de señales intracelulares. Esto incluye la
activación de canales de calcio en la membrana de los
osteocitos, permitiendo la entrada de calcio desde el
líquido intersticial circundante.
3. Transferencia de Calcio Intracelular: La entrada de calcio en los osteocitos provoca un aumento
significativo en la concentración de calcio dentro de estas células. Este aumento de calcio
intracelular es esencial para varias funciones celulares y señalización intracelular.
4. Reducción de Esclerostina: La PTH intermitente también reduce la producción de una proteína
llamada esclerostina por parte de los osteocitos. La esclerostina es una molécula inhibidora de la
formación ósea. Al reducir la esclerostina, la PTH estimula la diferenciación osteoblástica, lo que
significa que promueve la transformación de células precursoras en osteoblastos maduros. Además,
la PTH inhibe la apoptosis (muerte celular programada) de los osteoblastos, lo que aumenta la
población de estas células involucradas en la formación de hueso.
5. Estimulación Indirecta de la Síntesis Ósea: La reabsorción osteoclástica, que es la acción de los
osteoclastos en la degradación del hueso, libera factores de crecimiento almacenados en la matriz
ósea. Entre estos factores se encuentran el factor de crecimiento similar a la insulina 1 (IGF-1), el
IGF-2 y el factor de crecimiento fibroblástico (FGF). Estos factores de crecimiento son liberados
durante la reabsorción ósea y quedan atrapados en la matriz. La PTH, al estimular la resorción ósea,
contribuye a liberar estos factores, que a su vez estimulan la síntesis ósea.
6. Comunicación Osteocito-Osteoblasto: Los osteocitos no solo retienen el calcio para sí mismos, sino
que también establecen una comunicación bidireccional con los osteoblastos, células encargadas de
la formación de hueso nuevo. Esta comunicación se logra a través de complejas estructuras
conocidas como uniones en hendidura, permitiendo la transferencia de calcio desde los osteocitos a
los osteoblastos.
7. Bombeo de Calcio a la Matriz Extracelular: Los osteoblastos, que han recibido calcio de los
osteocitos, bombean activamente este calcio hacia la matriz extracelular del hueso. En la matriz
extracelular, el calcio se combina con el fosfato para formar cristales de hidroxiapatita, el
componente mineral principal del hueso.
8. Mineralización Ósea: La mineralización ósea es el proceso en el cual estos cristales de hidroxiapatita
se depositan en la matriz ósea, fortaleciendo y endureciendo el hueso. Este proceso es fundamental
para mantener la densidad ósea y la resistencia del esqueleto.
5.6.- Hipercalcemia hipocalciúrica familiar
La HHF es un trastorno genético hereditario que afecta a la regulación del calcio en el cuerpo.
1. Antecedentes Genéticos: La causa principal de la HHF es una mutación en el gen que codifica el
receptor de calcio sensible (CaSR) presente en las células paratiroideas y en los riñones por lo que
los pacientes con HHF presentan concentraciones plasmáticas de calcio entre un 10% y un 30% por
encima de los valores normales, suelen ser asintomáticos o presentan síntomas leves, lo que contrasta
con el nivel de hipercalcemia observado. La paradoja de la HHF radica en que, a pesar de los
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
elevados niveles de calcio plasmático, la concentración de la hormona paratiroidea (PTH) en estos
pacientes suele mantenerse dentro de los límites normales. Esto sugiere una regulación intacta de la
secreción de PTH, pero con un umbral ajustado a una mayor concentración de calcio plasmático.
2. Función Normal del CaSR: El CaSR es un receptor que actúa como un "sensor" de los niveles de
calcio en el organismo. Está especialmente presente en las glándulas paratiroides (pequeñas
glándulas ubicadas en el cuello) y en los túbulos renales de los riñones. Su función es detectar los
cambios en los niveles de calcio en la sangre y regular la secreción de la hormona paratiroidea (PTH)
en consecuencia. Además de las glándulas paratiroides, los túbulos renales gruesos ascendentes y el
túbulo contorneado distal también poseen receptores de calcio en sus membranas basolaterales. Estos
receptores responden a las variaciones en la concentración de calcio plasmático y regulan la
reabsorción renal de calcio. Cuando estos receptores están mutados, la reabsorción de calcio en los
riñones no puede inhibirse hasta que la concentración de calcio plasmático alcance niveles
anormalmente altos, lo que resulta en una mayor reabsorción de calcio y en la característica
hipocalciuria observada en la HHF.
3. Regulación del Calcio: En una persona sana, cuando los niveles de calcio en la sangre son bajos, el
CaSR estimula la liberación de PTH por parte de las glándulas paratiroides. La PTH aumenta la
reabsorción de calcio en los riñones y la liberación de calcio desde el hueso, lo que eleva los niveles
de calcio en la sangre.
4. Mutación en el CaSR: En pacientes con HHF, existe una mutación en el gen que codifica el CaSR.
Esta mutación hace que el receptor sea menos sensible al calcio, lo que significa que no "detecta"
adecuadamente los niveles elevados de calcio en la sangre. Este trastorno, de herencia autosómica
dominante, se asocia con diversas mutaciones puntuales en el gen del receptor de calcio. En
individuos heterocigotos, que tienen una copia normal y una mutada del gen, coexisten ambos tipos
de receptores de calcio. Por otro lado, los individuos homocigotos, que poseen dos copias mutadas,
presentan una hipercalcemia más grave, hiperparatiroidismo severo y manifestaciones clínicas más
pronunciadas, como alteraciones neurológicas y desmineralización ósea, lo que puede resultar en un
cuadro potencialmente mortal, a menos que se realice la extirpación de las glándulas paratiroides.
5. Consecuencias de la Mutación: A pesar de tener niveles de calcio en sangre más altos de lo normal
(entre un diez y un treinta por ciento más), la mayoría de los pacientes con HHF no experimentan los
síntomas típicos de la hipercalcemia, como fatiga, confusión o cálculos renales. Esto se debe a que
sus células paratiroideas no liberan PTH en respuesta a la hipercalcemia, ya que el receptor mutado
no lo detecta.
6. Diagnóstico: La HHF se diagnostica mediante pruebas genéticas que identifican la mutación en el
gen del CaSR. También se pueden realizar análisis de sangre para confirmar los niveles elevados de
calcio en la sangre y la concentración de PTH normal o ligeramente elevada. Así pues, los niveles de
calcio en la orina estarán disminuidos.
7. Tratamiento: Dado que los pacientes con HHF generalmente no presentan síntomas graves debido a
su hipercalcemia, el tratamiento puede no ser necesario. Sin embargo, en casos graves o cuando se
desarrollan complicaciones, se pueden usar medicamentos para reducir los niveles de calcio en la
sangre (calciomiméticos que activen la PTH).
5.7.- Vitamina D
1. Metabolismo vitamina D.
El metabolismo de la vitamina D es un proceso complejo que involucra varias etapas y órganos en el cuerpo:
a. Síntesis Cutánea: La primera fuente de vitamina D es la síntesis cutánea. Cuando la piel está
expuesta a la luz solar ultravioleta B (UVB), una molécula precursora llamada 7-
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
dehidrocolesterol, presente en las células de la epidermis, se convierte en previtamina D3. Esta
previtamina D3 es un compuesto inactivo.
b. Conversión en Vitamina D3: La previtamina D3 es transportada a los capilares sanguíneos de la
dermis y epidermis, donde la radiación UVB la convierte en vitamina D3 (colecalciferol). Esta
forma es aun biológicamente inactiva y requiere más procesamiento.
c. Hígado: La vitamina D3 es transportada por la sangre al hígado, donde se convierte en 25-
hidroxivitamina D [25(OH)D], también conocida como calcidiol. Este paso es catalizado por una
enzima llamada 25-hidroxilasa, y la 25(OH)D es la forma circulante principal de la vitamina D
en el cuerpo.
d. Riñones:La 25(OH)D es transportada al riñón, donde se lleva a cabo una segunda hidroxilación,
catalizada por la 1α-hidroxilasa renal. Este paso convierte la 25(OH)D en 1,25-dihidroxivitamina
D [1,25(OH)2D], también conocida como calcitriol, que es la forma activa de la vitamina D.
2. Vitamina D y la reabsorción de calcio.
La absorción de calcio en el intestino es un proceso altamente regulado que implica varios pasos clave:
• Paso 1: Captación de Calcio. El calcio (Ca2+) ingresa a la célula intestinal a través de canales de
calcio TRPV6 en la membrana apical.
• Paso 2: Unión a Proteínas Ligantes. Una vez dentro de la célula, el Ca2+ se une a proteínas ligantes,
especialmente la calbindina. Estas proteínas de alta afinidad para el calcio ayudan a regular su
concentración intracelular.
• Paso 3: Tamponado y Mantenimiento de Gradiente. El calcio es tamponado eficazmente en el citosol
de la célula por las proteínas ligantes y se almacena en compartimentos como el retículo
endoplásmico rugoso (RER) y las mitocondrias. Esto mantiene un gradiente de concentración
favorable para que el calcio pueda entrar desde el interior de la célula hacia la membrana apical.
• Paso 4: Expulsión de Calcio. El calcio es expulsado de la célula intestinal hacia la circulación
sanguínea a través de la membrana basolateral. Este proceso de expulsión implica una bomba de
calcio (ATPasa de calcio) y un intercambiador de sodio y calcio (Na-Ca).
• Vitamina D y Efectos Genómicos. La vitamina D, especialmente en su forma activa llamada
calcitriol, desempeña un papel esencial en la absorción de calcio. La vitamina D actúa a nivel
genómico, estimulando la síntesis de canales y bombas de calcio en las células intestinales. También
induce la producción de proteínas de unión al calcio y otras proteínas, como la fosfatasa alcalina, que
contribuyen al proceso de absorción.
En conjunto, estos pasos permiten que el intestino absorba eficazmente el calcio de la dieta y lo haga
disponible para su uso en el cuerpo.
3. Vitamina D y la reabsorción de fosforo.
a. Captación Inicial de Fósforo. El fósforo se encuentra en el lumen del intestino delgado y debe ser
captado por las células epiteliales intestinales para su absorción en la circulación. Para lograr
esto, las células intestinales utilizan un transportador de sodio y fósforo llamado cotransportador
NaPi.
b. Acción de la Vitamina D. La vitamina D, específicamente en su forma activa conocida como
1,25-dihidroxivitamina D o calcitriol, desempeña un papel fundamental en este proceso. El
calcitriol actúa como un modulador de la síntesis de proteínas transportadoras de fósforo en las
células de la mucosa intestinal. Estimula la síntesis y expresión del cotransportador NaPi en la
membrana apical de las células epiteliales intestinales. Al aumentar la cantidad de
cotransportadores NaPi en la membrana apical, el calcitriol mejora la capacidad de las células
intestinales para captar fósforo desde el lumen intestinal.
c. Resultado: Mayor Absorción de Fósforo Como resultado de la acción de la vitamina D, se
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
aumenta la absorción de fósforo desde el intestino hacia las células de la mucosa intestinal. Este
proceso facilita la entrada de fósforo en el torrente sanguíneo y asegura un suministro adecuado
de fósforo en el organismo.
4. Déficit de vitamina D.
a. Raquitismo: El raquitismo es un trastorno metabólico del tejido óseo que afecta principalmente a
los niños en crecimiento. Se caracteriza por una mineralización inadecuada del hueso en
desarrollo y suele estar relacionado con una deficiencia de vitamina D. Aquí hay un análisis más
detallado:
i. Causas: La principal causa del raquitismo es la deficiencia de vitamina D, que es esencial
para la absorción intestinal de calcio y fósforo. La vitamina D se obtiene a través de la
exposición solar, la dieta y la síntesis cutánea. La falta de cualquiera de estos elementos
puede llevar a una deficiencia de vitamina D.
ii. Efectos en los huesos: La deficiencia de vitamina D resulta en una incapacidad para
absorber y utilizar adecuadamente el calcio y el fósforo, lo que conduce a la formación de
huesos blandos y deformidades óseas. Los huesos largos, como los de las piernas, pueden
volverse arqueados, y las muñecas pueden ensancharse. Además, el cartílago de
crecimiento no se mineraliza adecuadamente, lo que puede provocar retrasos en el
crecimiento.
iii. Síntomas adicionales: Los niños con raquitismo pueden experimentar dolor óseo,
debilidad muscular, retraso en el desarrollo motor y dental, y un mayor riesgo de
fracturas. En casos graves, los niveles bajos de calcio en sangre pueden llevar a
convulsiones.
b. Osteomalacia: La osteomalacia es una afección similar al raquitismo, pero que afecta a los
adultos. Aquí hay más detalles:
c. Causas: Al igual que en el raquitismo, la causa principal de la osteomalacia es la deficiencia de
vitamina D. También puede ser causada por trastornos que afectan la capacidad del riñón para
activar la vitamina D o problemas de absorción intestinal.
d. Efectos en los huesos: La insuficiencia de vitamina D y la mala absorción de calcio y fósforo
llevan a la formación de huesos con una mineralización inadecuada. Esto aumenta
significativamente la fragilidad de los huesos, lo que se traduce en un mayor riesgo de fracturas.
Los huesos pueden ser tan frágiles que actividades normales como caminar pueden causar
fracturas. Además, el tejido óseo mal mineralizado puede dar lugar a deformidades óseas en
adultos.
e. Síntomas adicionales: Los pacientes con osteomalacia pueden experimentar dolor óseo
generalizado, debilidad muscular, fatiga, espasmos musculares involuntarios y parestesias debido
a la hipocalcemia y la irritabilidad neuromuscular.
6.- CALCITONINA.
La calcitonina es una hormona peptídica de 32
aminoácidos que se origina en las células C,
también conocidas como células parafoliculares,
de la glándula tiroides. A pesar de su localización
en la tiroides, la función principal de la calcitonina
radica en la regulación del metabolismo mineral y
la renovación ósea, lo que hace que su relación con
las funciones principales de la tiroides sea algo
fortuito.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
La síntesis de calcitonina ocurre mediante la vía secretora, que implica el procesamiento postraducción de
una procalcitonina más grande. La escisión alternativa de este producto de gen de calcitonina resulta en
varios péptidos con actividad biológica, pero en las células C, la calcitonina es el único péptido sintetizado
en cantidades biológicamente significativas.
La liberación de calcitonina se activa cuando la concentración extracelular de calcio aumenta por encima de
lo normal. En respuesta a esto, la calcitonina reduce la concentración de calcio en la sangre. Sin embargo, es
importante señalar que la calcitonina parece tener un papel limitado en la regulación a corto plazo de la
concentración plasmática de calcio en humanos, ya que la extirpación total de la tiroides no afecta
significativamente esta concentración, siempre que las glándulas paratiroides, que también regulan el calcio,
estén intactas.
A pesar de esta aparente falta de importancia en la homeostasis del calcio en humanos, la calcitonina
desempeña un papel crítico en muchas especies no mamíferas. Por ejemplo, en peces teleósteos, donde la
concentración de calcio en el agua marina es relativamente alta, la calcitonina se secreta en respuesta al
aumento de la concentración plasmática de calcio y reduce la reabsorción ósea, restableciendo así la
concentración plasmática de calcio a niveles normales. La calcitonina del salmón, que es diferente de la
humana en varios aminoácidos, es aún más potente en la inhibición de la función de los osteoclastos
humanos.
La acción principal de la calcitonina se ejerce a través de receptores de calcitonina, que son receptores
acoplados a proteína G (GPCR). Estos receptores pueden activar la adenilato ciclasa o la fosfolipasa C,
dependiendo del tipo de célula diana. En el hueso, la calcitonina actúa principalmente sobre los osteoclastos,
inhibiendo su actividad y reduciendo la velocidad de renovación ósea. También reduce la osteólisis
osteocitaria, lo que, junto con su efecto sobre los osteoclastos, provoca una disminución de la concentración
de calcio en la sangre, lo que se conoce como hipocalcemia, cuando se administra en dosis farmacológicas.
A pesar de su efectividad en la regulación del calcio, el uso clínico de la calcitonina es limitado debido a la
desensibilización de los receptores de calcitonina que ocurre después de varias horas de exposición a altas
concentraciones de la hormona. Además, la calcitonina también tiene efectos en el riñón, donde induce una
leve fosfaturia, natriuresis y calciuresis, contribuyendo así a sus efectos hipocalcémicos e hipofosfatémicos
agudos. Sin embargo, estos efectos renales son de corta duración y no desempeñan un papel importante en la
regulación renal general del calcio, el fósforo o el sodio.
7.- ROL DE LOS ESTEROIDES SEXUALES EN LA FORMACIÓN DE HUESOS.
Los esteroides sexuales, como la testosterona en los varones y el estradiol en las mujeres, desempeñan un
papel fundamental en el mantenimiento de una masa ósea normal. La disminución de los niveles de estradiol
después de la menopausia en las mujeres aumenta el riesgo de osteoporosis, caracterizada por una
disminución en la masa de hueso cortical y trabecular debido a la reducción de la matriz ósea. En los
varones, la osteoporosis es menos común debido a su mayor masa ósea durante la vida adulta y a la
disminución gradual de los niveles de testosterona con la edad, en contraste con la disminución abrupta de
estradiol en las mujeres durante la menopausia.
Los glucocorticoides también ejercen un efecto sobre la masa ósea, siendo más evidente en situaciones de
exceso de estos esteroides, lo que puede provocar osteoporosis. Esto se relaciona con los efectos de los
glucocorticoides en la producción de OPG (osteoprotegerina) y RANKL (receptor activador del factor
nuclear kappa-B ligando).
Aunque aún no se han definido completamente los mecanismos celulares específicos que median en la
acción de los andrógenos, estrógenos y glucocorticoides sobre el hueso, es importante destacar que, a pesar
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
de la pérdida ósea asociada con la deficiencia de andrógenos o estrógenos o el exceso de glucocorticoides, se
mantiene el equilibrio entre la síntesis y la degradación ósea. En otras palabras, se sigue formando nuevo
hueso en los lugares donde se había reabsorbido el hueso anteriormente, y no se produce una acumulación
excesiva de osteoide no mineralizado. Sin embargo, la cantidad de nuevo hueso formado en cualquier punto
es menor que la cantidad de hueso reabsorbido, lo que conduce a una disminución general de la masa ósea
debido a un cambio cuantitativo en el equilibrio entre formación y reabsorción ósea que afecta a varios
lugares en el cuerpo.
8.- EJE HIPOTÁLAMO-HIPÓFISIS-ADRENAL.
Las dos glándulas suprarrenales, con un peso aproximado de
4 g cada una, se hallan en los polos superiores de los
riñones. Se compone de dos porciones diferentes, la médula
suprarrenal y la corteza suprarrenal. La médula suprarrenal,
que ocupa el 20% central de la glándula, se relaciona desde
el punto de vista funcional con el sistema nervioso
simpático; secreta las hormonas adrenalina y noradrenalina
en respuesta a la estimulación simpática. A su vez, estas hormonas provocan casi los mismos efectos que la
estimulación directa de los nervios simpáticos en todas las regiones del cuerpo. La corteza suprarrenal
secreta un grupo completamente diferente de hormonas, llamadas corticoesteroides. Todas estas hormonas se
sintetizan a partir del esteroide colesterol y todas poseen una fórmula química parecida. Sin embargo, las
pequeñas variaciones de su estructura molecular proporcionan diferencias funcionales muy importantes.
La corteza suprarrenal secreta los dos tipos principales de hormonas corticosuprarrenales, los
mineralocorticoides y los glucocorticoides. Además, produce pequeñas cantidades de hormonas sexuales, en
particular de andrógenos, que inducen los mismos efectos que la hormona sexual masculina testosterona. En
general, son de escasa importancia, pero cuando se secretan en grandes proporciones en algunos trastornos
de la corteza suprarrenal, causan los efectos virilizantes consiguientes. Los mineralocorticoides reciben este
nombre porque afectan sobre todo a los electrólitos (los «minerales») del compartimiento extracelular,
especialmente al sodio y al potasio. Los glucocorticoides se denominan así porque poseen efectos
importantes de aumento de la glucemia. Además, influyen en el metabolismo de las proteínas y de los
lípidos. Los más importantes son: la aldosterona, que es el mineralocorticoide principal, y el cortisol, que es
el glucocorticoide principal.
Esta corteza tiene 3 capas diferentes:
• Zona glomerular (15%), capa delgada de células
situada inmediatamente por debajo de la cápsula. Estas células
son las únicas de la glándula suprarrenal capaces de secretar
cantidades importantes de aldosterona porque contienen la
enzima aldosterona sintetasa, necesaria para la síntesis de la
hormona. Secreción controlada sobre todo por las
concentraciones de angiotensina II y la concentración de potasio
en el líquido extracelular.
• Zona fascicular (75%), capa media y más ancha
secreta los glucocorticoides cortisol y corticosterona, así como
pequeñas cantidades de andrógenos y estrógenos suprarrenales. La secreción de estas células está
controlada, en gran parte, por el eje hipotálamo-hipofisario a través de la corticotropina (ACTH).
• Zona reticular, la capa más profunda de la corteza, secreta los andrógenos suprarrenales
dehidroepiandrosterona (DHEA) y androstenediona. La ACTH también regula la secreción de estas
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
células, aunque en ella pueden intervenir otros factores tales como la hormona corticótropa
estimuladora de los andrógenos, liberada por la hipófisis.
Todas las hormonas esteroideas humanas, incluidas las producidas por la corteza suprarrenal, se sintetizan a
partir del colesterol, Si bien las células de la corteza suprarrenal pueden sintetizar de novo pequeñas
cantidades de colesterol a partir del acetato, casi el 80% del colesterol empleado para la síntesis de
esteroides proviene de las lipoproteínas de baja densidad (LDL) del plasma circulante. Las LDL, que
transportan altas concentraciones de colesterol, difunden desde el plasma al líquido intersticial para unirse a
receptores específicos localizados en estructuras de la membrana de la célula corticosuprarrenal conocidas
como depresiones revestidas. Estas depresiones penetran en el citoplasma por endocitosis, transformándose
en vesículas que, por último, se fusionan con los lisosomas y liberan el colesterol destinado a la síntesis de
los esteroides suprarrenales. El transporte del colesterol a las células suprarrenales está sometido a
mecanismos de retroalimentación que pueden modificar en gran medida la cantidad disponible para la
síntesis de esteroides. Por ejemplo, la ACTH, que estimula la síntesis de esteroides suprarrenales, incrementa
el número de receptores de LDL de la célula corticosuprarrenal y la actividad de las enzimas que liberan el
colesterol a partir de las LDL. Cuando el colesterol entra en la célula, pasa a las mitocondrias, donde se
escinde por acción de la enzima colesterol desmolasa para formar pregnenolona; este es el paso que acaba
limitando la síntesis de los esteroides suprarrenales. Este paso inicial de la síntesis de los esteroides en las
tres zonas de la corteza suprarrenal se estimula por los diversos factores que controlan la secreción de los
principales productos hormonales, aldosterona y cortisol. Casi todas estas etapas suceden en dos orgánulos
celulares, las mitocondrias y el retículo endoplásmico, pero algunas tienen lugar en las primeras y otras en el
segundo. Destacamos las siguientes etapas:
Mencionar que la 17α-hidroxilasa no
abunda en la capa glomerular. De
este modo, la capa fascicular y, en
mucha menor medida, la reticular
sintetizan el cortisol.
Tras la síntesis las hormonas
corticosuprarrenales se unen a
proteínas del plasma; alrededor del
90% del cortisol se transporta unido
a la globulina de unión a corticoides
(CBG), también conocida como
transcortina, cuya afinidad por el
cortisol es 30 veces mayor que por la
aldosterona. Otro 7% del cortisol
circulante se encuentra unido a la
albúmina. De este modo, solo el 3-
4% del cortisol circulante se
encuentra en forma libre. Esta unión
tan fuerte a las proteínas del plasma
reduce la velocidad de eliminación
del cortisol plasmático; por tanto, el
cortisol posee una semivida
relativamente larga, de 60 a 90 min.
Tan solo el 60% de la aldosterona
circulante se une a las proteínas del plasma, de modo que el 40% queda en forma libre; en consecuencia, su
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
semivida es bastante reducida (20min).
El cortisol, una vez cumplió sus funciones, se transforma en cortisona en un primer paso (reversible). Este
proceso de conversión a cortisona ocurre mediante la acción de una de las dos isoformas de la enzima 11β-
hidroxiesteroide deshidrogenasa (11β-HSD). La isoforma específica llamada 11β-HSD2, que se encuentra en
los túbulos distales y colectores del riñón, participa en un segundo paso. Aquí, cataliza una conversión
irreversible de cortisol a cortisona en el riñón.
8.1.- Glucocorticoides: Colesterol
Síntesis y secreción de colesterol.
1. Inicio del Proceso. La liberación de la hormona liberadora de corticotropina (CRH)
por neuronas hipotalámicas inicia la regulación de cortisol. Esto ocurre en el ritmo
circadiano normal o como respuesta al estrés.
2. Acciones de la CRH. La CRH llega a la hipófisis anterior y se une al receptor CRH-
R1. Este receptor es un GPCR (proteína acoplada a la membrana por una proteína
G). La unión activa la Gαs, que estimula la adenilato-ciclasa, aumenta AMPc, y
estimula la liberación de ACTH.
3. Secreción de ACTH. La ACTH estimula la síntesis y secreción de cortisol en las
capas fasciculares suprarrenales. El cortisol ejerce retroalimentación negativa sobre
la liberación de CRH.
a. Otros Reguladores. Además de la CRH, los núcleos paraventriculares producen hormona
antidiurética (ADH), que también influye en la ACTH; la ADH es un potente secretagogo de
la ACTH.
4. Interacción con MC2R. En la corteza suprarrenal, la ACTH se une
al receptor MC2R en las células secretoras de esteroides. Sin embargo, dado
que solo las células de las capas fascicular y reticular poseen la 17α-
hidroxilasa necesaria para sintetizar cortisol, estas células son las únicas que
secretan cortisol en respuesta a la ACTH.
5. Señalización Celular: MC2R se acopla a una proteína G y estimula
la adenilato-ciclasa que produce un aumento de AMPc el cual activa la
proteína-cinasa A (PKA).
6. Efectos de la ACTH:
a. La ACTH estimula rápidamente la conversión de colesterol a
pregnenolona mediante la enzima esteroide desmolasa.
b. A largo plazo, aumenta la síntesis de varias proteínas necesarias
para la síntesis de cortisol:
i. Enzimas P-450 involucradas en la síntesis de cortisol.
ii. Receptor de LDL necesario para la captación de
colesterol circulante.
iii. 3-hidroxi-3-metilglutaril coenzima A (HMG-CoA)
reductasa, limitante en la síntesis de colesterol.
7. Atrofia sin ACTH. La falta de ACTH provoca la atrofia de las capas
fascicular y reticular de la corteza suprarrenal.
El cortisol ejerce un control mediante retroalimentación negativa sobre el propio eje que estimula su
secreción.
• Retroalimentación sobre la hipófisis anterior:
En las células corticotropas de la hipófisis anterior, el cortisol se une a un receptor en el citosol. Después de
esta unión, el complejo se traslada al núcleo, donde se une a los Elementos de Respuesta a Glucocorticoides
(GRE), regulando así la expresión génica. El cortisol inhibe la síntesis del receptor de la hormona liberadora
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
de corticotropina (CRH) y de la hormona adrenocorticotropa (ACTH). Además, altos niveles de cortisol
inhiben la liberación de ACTH previamente sintetizada y almacenada en vesículas.
• Retroalimentación sobre el hipotálamo:
La retroalimentación negativa del cortisol sobre las neuronas hipotalámicas secretoras de CRH es menos
significativa que en las células corticotropas. El cortisol plasmático reduce la expresión génica de los
péptidos de CRH en las neuronas paraventriculares del hipotálamo. También inhibe la liberación de CRH ya
sintetizada. Los glucocorticoides sintéticos tienen un efecto similar.
• Control superior por el SNC:
o Naturaleza circadiana y pulsátil de la secreción: La liberación de ACTH y cortisol sigue un
ritmo circadiano, es decir, un patrón que se repite aproximadamente cada 24 horas. El núcleo
supraquiasmático del hipotálamo, que recibe información de la retina, desempeña un papel
esencial en la regulación de los ritmos circadianos del cuerpo. La actividad secretora de
ACTH alcanza su punto máximo por la mañana y disminuye hacia la tarde y noche. En
personas con pérdida de la función visual, como aquellas ciegas, se observa una pérdida de
los ritmos circadianos.
o Además, el SNC responde a factores estresantes, tanto físicos como psicológicos y
bioquímicos. En situaciones de estrés, hay un aumento en la secreción de CRH y, por ende,
de ACTH. Por ejemplo, el estrés bioquímico, como la hipoglucemia, estimula la liberación de
CRH y ACTH, lo que resulta en un aumento del cortisol para elevar los niveles de glucosa en
sangre.
Acciones de los glucocorticoides.
Casi todas las acciones mejor conocidas de los glucocorticoides se deben a sus acciones genómicas que
influyen (ya sea de forma positiva o negativa) en la transcripción de una serie de genes mediante los
elementos de respuesta a glucocorticoides. Sin embargo, los glucocorticoides también pueden ejercer
acciones no genómicas que se producen en un periodo breve de tiempo (entre 0 y 3 horas) y que no se ven
inhibidas por el bloqueo de la transcripción génica.
El cortisol, como otras hormonas esteroideas, ejerce sus efectos mediante su interacción inicial con los
receptores intracelulares de las células efectoras. El cortisol es liposoluble y difunde con facilidad a través
de la membrana celular. Una vez en el interior de la célula, se une con el receptor de glucocorticoides (GR)
que mayoritariamente se encuentra en el citoplasma, donde, en su forma libre, establece un complejo con
una proteína chaperona; cuando el cortisol se une al GR, la chaperona se separa del complejo, permitiendo
que el complejo cortisol-GR se traslade al núcleo celular. En este sitio, el complejo cortisol-receptor se
asocia con los Elementos de Respuesta a Glucocorticoides (GRE) en el extremo 5' no traducido de múltiples
genes, lo que puede resultar en la regulación al alza o a la baja de su expresión.
Los efectos más destacables de los glucocorticoides son:
• Síntesis Hepática de Glucosa y Glucógeno (Hígado). Los glucocorticoides inducen enzimas
hepáticas que participan en la gluconeogénesis y el metabolismo de aminoácidos. Este proceso es
crucial para la producción de glucosa en situaciones de ayuno prolongado o estrés, proporcionando
un suministro constante de glucosa al torrente sanguíneo.
• Degradación de Proteínas Musculares (Músculo). La estimulación de la degradación de proteínas
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
musculares por los glucocorticoides tiene lugar en condiciones de estrés o necesidad energética. La
liberación de aminoácidos resultantes se dirige al hígado para la gluconeogénesis o se utiliza como
fuente de energía.
• Lipólisis y Movilización de Grasa (Tejido Adiposo). La lipólisis inducida por glucocorticoides libera
ácidos grasos del tejido adiposo. Esta respuesta es esencial para proporcionar energía adicional,
particularmente en situaciones de estrés, y es parte de la adaptación del organismo a la movilización
de recursos.
• Uso Clínico en Trastornos Inflamatorios. En trastornos inflamatorios como vasculitis y artritis, los
glucocorticoides actúan como potentes antiinflamatorios y suprimen respuestas inmunológicas. Su
uso en neoplasias malignas se centra en la reducción de la inflamación y la respuesta inmune
exacerbada.
• Impacto en el Sistema Óseo. Los glucocorticoides afectan negativamente la salud ósea al interferir
con la formación de nuevo hueso por los osteoblastos y al reducir la absorción de calcio en el tracto
digestivo. Este proceso a largo plazo puede llevar a la osteoporosis.
• Efectos sobre el Sistema Nervioso Central (SNC). Los glucocorticoides tienen un impacto en el
estado de ánimo y la cognición. Su uso prolongado puede contribuir a efectos secundarios
neuropsiquiátricos, incluyendo cambios en la percepción y en la función cognitiva.
En resumen, los glucocorticoides desempeñan un papel vital en la adaptación del organismo a condiciones
de estrés y desafío fisiológico. Sus efectos beneficiosos en la regulación metabólica y la respuesta
antiinflamatoria son fundamentales, pero su uso prolongado o desregulado puede tener consecuencias
adversas, subrayando la importancia de su administración clínica cuidadosa.
Desregulación de los glucocorticoides.
• Síndrome de Cushing/ Hiperfunción corticosuprarrenal. Trastorno producido por exceso
glucocorticoides: cortisol. Causas:
o Adenomas hipofisiarios (secundario) o suprarrenales (primario)
o Tumores malignos éxtrahipofisiarios: Tumor en otra parte del cuerpo que produce hormona
liberadora de adrenocorticotropina
o Uso continuado de tratamiento glucocorticoides: Cushing exógeno
o Manifestaciones:
▪ Hiperglucemia. Aunque la función principal del cortisol es disminuir la glucemia, en
la enfermedad de Cushing, hay una resistencia a los efectos de la insulina debido al
exceso crónico de cortisol. Esto conduce a niveles elevados de glucosa en sangre.
▪ Hipertensión. El aumento de la aldosterona, que es un mineralocorticoide liberado en
exceso en la enfermedad de Cushing, contribuye a retención de sodio y líquidos, lo
que eleva la presión arterial.
▪ Hipervolemia. La aldosterona, aumentada en la enfermedad de Cushing, promueve la
retención de sodio y agua, resultando en un aumento del volumen sanguíneo y la
presión arterial.
▪ Obesidad/Cara luna llena/Jiba de búfalo. El patrón de distribución centrípeta de la
grasa, especialmente en la región del tronco y la cara, se debe al efecto del exceso de
cortisol, que favorece la acumulación de grasa en estas áreas específicas.
▪ Estrías purpúreas/equimosis. El aumento del metabolismo de las proteínas y la
hiperglucemia resultante contribuyen a la pérdida de masa muscular y a la fragilidad
de la piel, manifestándose como estrías purpúreas (líneas rojas en la piel) y mayor
propensión a las equimosis (moretones).
▪ Hipopotasemia. La sobreproducción de aldosterona conduce a una excesiva retención
de sodio y excreción de potasio, provocando niveles bajos de potasio en sangre.
▪ Hirsutismo. El desequilibrio hormonal en la enfermedad de Cushing puede llevar al
desarrollo excesivo de vello en áreas donde normalmente no crece en cantidades
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
significativas.
▪ Osteoporosis/Debilidad. El aumento del metabolismo de las proteínas puede
contribuir a la pérdida de masa ósea, aumentando el riesgo de osteoporosis y debilidad
muscular.
▪ Trastornos de Ánimo. Las fluctuaciones en los niveles de cortisol y otros
desequilibrios hormonales pueden tener un impacto en el estado de ánimo,
contribuyendo a cambios emocionales frecuentes.
• Hipofunción corticosuprarrenal (insuficiencia corticosuprarrenal): enfermedad de Addison
o Causas. La enfermedad de Addison se debe a la incapacidad de la corteza suprarrenal para
fabricar suficientes hormonas corticales; a su vez, en un elevado número de casos, la causa
obedece a una atrofia o lesión primaria de la corteza suprarrenal, estas casusas primarias se
ban en: autoinmune (80% de los casos), tumor, hemorragia, infecciosa o suprarrenalectomía
bilateral. Así pues, esta patología también puede ser secundaria a un deterioro en la función
de la hipófisis, que no consigue producir suficiente ACTH. Cuando la producción de ACTH
es demasiado baja, la de cortisol y aldosterona disminuye y, finalmente, las glándulas
suprarrenales pueden llegar a atrofiarse debido a la ausencia de estimulación de ACTH. La
insuficiencia suprarrenal secundaria es mucho más común que la enfermedad de Addison,
que en ocasiones se denomina insuficiencia suprarrenal primaria.
o Manifestaciones:
▪ Déficit de Aldosterona. La falta de aldosterona afecta la capacidad de los riñones para
regular los electrolitos, lo que lleva a una disminución del sodio y un aumento del
potasio. Esto contribuye a la hipovolemia (baja cantidad de sangre circulante),
deshidratación y hipotensión ortostática
▪ Reducción de Gluconeogénesis Hepática. Hipoglucemia. La insuficiencia adrenal
reduce la capacidad del hígado para producir glucosa a partir de fuentes no glucídicas
(gluconeogénesis). La hipoglucemia resultante puede causar debilidad y fatiga debido
a la falta de glucosa, que es esencial para la energía celular.
▪ Aumento de ACTH y MSH (hormona estimulante de los menalocitos.
Manifestaciones Cutáneas y Mucosas:
• Pigmentación de Mucosas: La melanina se deposita en las mucosas, como las
encías y el revestimiento oral, causando un oscurecimiento.
• Hiperpigmentación de Pezones y Zonas de Fricción: Las áreas de fricción de
la piel y los pezones pueden volverse más oscuros debido al aumento de la
producción de melanocitos.
• Hiperpigmentación de Piel: La piel en general puede volverse más oscura
debido al aumento de la síntesis de melanina.
• Pecas: La sobreproducción de melanocitos puede dar lugar a la formación de
pecas.
• Oscurecimiento del Cabello: El cabello puede volverse más oscuro.
▪ Hiperpotasemia. Debilidad Muscular y Astenia Intensa. El aumento del potasio en
sangre puede afectar la excitabilidad de las células musculares, resultando en
debilidad muscular y fatiga intensa (astenia).
▪ Disminución de Vello
▪ Anorexia/Pérdida de Peso: La debilidad muscular y la fatiga pueden contribuir a la
pérdida de apetito (anorexia) y, como resultado, a la pérdida de peso.
▪ Intolerancia al Estrés. La falta de hormonas como el cortisol y la aldosterona afecta la
capacidad del cuerpo para manejar el estrés físico. Se produce una respuesta
inadecuada al estrés, llevando a la hipovolemia, deshidratación e hipotensión
ortostática.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Deficiencia de 21α-hidroxilasa.
La deficiencia de esta enzima conlleva una producción inadecuada tanto de glucocorticoides como de
mineralocorticoides, los niños afectados presentan síntomas de «pérdida salina» (hipotensión,
deshidratación) y de ausencia de glucocorticoides (hipoglucemia).
El cuerpo reacciona de forma natural intentando compensar esta deficiencia mediante el aumento de la
secreción de ACTH, que estimula la síntesis de cortisol y aldosterona. La ACTH también hace que la
glándula suprarrenal aumente de tamaño. Sin embargo, si la enzima mutada es completamente inactiva, no
tendrá lugar la síntesis de cortisol ni aldosterona, aunque las demás enzimas de la vía dedicada a la síntesis
de glucocorticoides y mineralocorticoides se expresarán en exceso. La consecuencia será una actividad
mayor de lo normal de la SCC, la 3β-HSD y la 17α-hidroxilasa, y el efecto neto será el aumento de la
síntesis tanto de moléculas precursoras como de andrógenos suprarrenales. La combinación de una
producción insuficiente de glucocorticoides y mineralocorticoides, una producción excesiva de andrógenos y
el mayor tamaño de la glándula suprarrenal da lugar al clásico síndrome clínico de la hiperplasia
suprarrenal congénita virilizante con pérdida salina. En las niñas, la presencia de un exceso de
andrógenos suprarrenales en el útero materno da lugar a unos genitales ambiguos al nacer, lo que debería dar
al pediatra un indicio del posible diagnóstico. En los hombres no existe ningún indicio equivalente, por lo
que las principales manifestaciones serán deshidratación, hipotensión e hiperpotasemia.
8.2.- Aldosterona.
Principal mineralocorticoide que desempeña un papel fundamental en la regulación del equilibrio
electrolítico y del volumen de fluidos en el organismo.
Síntesis de Aldosterona
1. Producción de Pregnenolona: La primera etapa es catalizada por la enzima citocromo P-450 (P-
450SCC), también conocida como esteroide colesterol side-chain cleavage (SCC). Esta enzima
convierte el colesterol en pregnenolona, actuando como el paso limitante en la velocidad de la
síntesis de hormonas esteroides.
2. Formación de Progesterona: La pregnenolona se somete a oxidación por la enzima 3β-
hidroxiesteroide deshidrogenasa (3β-HSD) en el retículo endoplásmico liso (REL), produciendo
progesterona.
3. Hidroxilación en la Posición 21: A diferencia de la síntesis de cortisol, las células glomerulares
utilizan una 21α-hidroxilasa (P-450c21) del REL para hidroxilar la progesterona en la posición 21,
generando 11-desoxicorticosterona (DOC).
4. Hidroxilación en las Mitocondrias: La 11β-hidroxilasa (P-450c11), presente en las mitocondrias,
añade un grupo ─OH en la posición 11 de la DOC, dando lugar a la formación de corticosterona.
Esta enzima realiza las mismas hidroxilaciones que en la síntesis de cortisol.
5. Aldosterona Sintasa (P-450aldo): Las células glomerulares, de manera exclusiva, contienen la
aldosterona sintasa (P-450aldo). Esta enzima, también conocida como 18-metiloxidasa, realiza dos
pasos cruciales: primero, añade un grupo ─OH al metilo de la posición 18, y luego oxida este
hidroxilo a un grupo aldehído, dando como resultado la aldosterona.
Regulación de la Secreción de Aldosterona
A diferencia del cortisol, la aldosterona no cuenta con depósitos previos en las células glomerulares de la
corteza suprarrenal, lo que implica que su liberación está estrictamente vinculada a la velocidad de síntesis
celular. La regulación de la secreción de aldosterona se ve influenciada por diversos factores fisiológicos,
siendo clave la concentración extracelular de potasio (K+) y la acción de la angiotensina II (ANG II).
1. Estimulación por ACTH:
a. Aunque la ACTH también participa en el estímulo de la síntesis de aldosterona en las células
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
glomerulares, su contribución fisiológica es eclipsada por otros reguladores más prominentes.
2. Angiotensina II y Potasio (K+):
a. La elevación de la concentración extracelular de K+ y la presencia de ANG II desempeñan
roles cruciales en el control de la secreción de aldosterona. Estos actúan aumentando la
actividad de las enzimas responsables de las etapas limitantes de la velocidad en la síntesis de
aldosterona, como la SCC y la aldosterona-sintasa.
3. Secreción y Unión en la Sangre:
a. Tras su secreción, aproximadamente el 37% de la aldosterona circula libremente en la sangre.
El restante se une de manera débil a la globulina fijadora de corticosteroides (CBG) en un
21%, o a la albúmina en un 42%.
Funciones de la aldosterona
La acción principal de la aldosterona es estimular al riñón para que reabsorba Na+ y agua y aumente la
excreción de K+. La aldosterona tiene unas acciones parecidas sobre el transporte de sodio y agua en el
colon, las glándulas salivales y las glándulas sudoríparas.
La aldosterona ejerce sus efectos a través de receptores de baja y alta afinidad. Mientras los receptores de
baja afinidad son análogos al receptor de glucocorticoides (GR), los receptores de alta afinidad, específicos
para mineralocorticoides (MR), están presentes en tejidos diana. El cortisol suele circular a concentraciones
muy superiores a las de la aldosterona (5-20 µg/dl frente a 2-8 ng/dl), es de esperar que el efecto biológico
de la aldosterona sobre cualquier diana posible se vea eclipsado por el del cortisol. Sin embargo, las células
que son diana de la aldosterona (sobre todo al comienzo del túbulo colector y en el túbulo colector cortical
del riñón) contienen 11β-HSD2, que convierte el cortisol en cortisona, un esteroide con una afinidad muy
baja por el MR. A diferencia de la 11β-HSD1, que convierte de forma intercambiable y reversible la
cortisona y el cortisol, la 11β-HSD2 no puede convertir la cortisona de nuevo en cortisol. A consecuencia de
ello, a nivel de la célula diana, la proporción entre cortisol y aldosterona es mucho menor que en la sangre,
donde predomina el cortisol.
La aldosterona, como el cortisol y las demás hormonas esteroideas, actúa principalmente modulando la
transcripción génica. En el riñón, la aldosterona induce una serie de eventos moleculares precisos. Aumenta
la transcripción de la bomba Na-K, favoreciendo la reabsorción distal de Na+. Además, potencia la
expresión de canales de Na+ apicales y de un cotransportador Na/K/Cl, contribuyendo al aumento neto de la
reabsorción de Na+ y excreción de K+. Desbalances en la acción de la aldosterona pueden tener
consecuencias clínicas significativas. Una deficiencia puede llevar a la retención de potasio, resultando en
hiperpotasemia, que puede ser potencialmente letal. Por otro lado, un exceso de aldosterona puede causar
pérdida excesiva de potasio, llevando a hipopotasemia e hipertensión.
Mecanismo celular de la acción de la aldosterona
La acción de la aldosterona en el transporte de sodio y potasio a través de las
células tubulares renales involucra varios pasos:
1. Difusión de Aldosterona. La aldosterona, siendo liposoluble en las
membranas celulares, difunde rápidamente hacia el interior de las células del
epitelio tubular.
2. Unión al Receptor Mineralocorticoide. La aldosterona se une al
receptor mineralocorticoide, una proteína configurada de manera que solo la
aldosterona o compuestos muy similares pueden unirse a ella. Aunque
también tiene afinidad por el cortisol, la enzima 11β-hidroxiesteroide
deshidrogenasa tipo 2 convierte la mayoría del cortisol en cortisona, que no
se une eficazmente al receptor.
3. Traslado al Núcleo y Síntesis de ARN Mensajero. El complejo
aldosterona-receptor o sus productos se trasladan al núcleo celular, donde
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
inducen la síntesis de ARN mensajero relacionado con el transporte de sodio y potasio.
4. Producción de Proteínas en el Citoplasma. El ARN mensajero se traslada al citoplasma, donde, junto
con los ribosomas, promueve la formación de proteínas. Estas proteínas incluyen enzimas y proteínas
transportadoras de membrana, esenciales para el transporte de sodio, potasio e hidrógeno a través de
la membrana celular.
5. Efecto en el Transporte Iónico. La trifosfatasa de adenosina de sodio-potasio aumenta
significativamente y actúa como componente principal de la bomba de intercambio de sodio y
potasio en la membrana basolateral de las células tubulares renales. Otras proteínas, como el canal de
sodio epitelial (ENaC), facilitan la difusión rápida de iones de sodio desde la luz del túbulo hacia la
célula. La bomba de sodio-potasio en la membrana basolateral bombea el sodio para completar el
proceso.
En resumen, la aldosterona no tiene un efecto inmediato sobre el transporte de sodio; su acción se
desencadena por una secuencia de eventos que culminan en la síntesis de las sustancias intracelulares
necesarias para el transporte iónico. Este proceso lleva aproximadamente 30 minutos para la producción de
nuevo ARN y alrededor de 45 minutos para alcanzar el efecto máximo en el transporte de sodio, siendo este
último observable después de unas horas.
Secreción de la aldosterona.
En el control de la síntesis de aldosterona por las células glomerulares de la corteza suprarrenal, tres
secretagogos desempeñan un papel fundamental:
• Angiotensina II (ANG II): Este es el secretagogo más importante. Se genera en la cascada del
sistema renina-angiotensina. La ANG II tiene una influencia significativa en la secreción de
aldosterona y desempeña un papel crucial en la regulación del equilibrio de electrolitos y la presión
arterial.
• Concentración Plasmática de Potasio (K+): Un aumento en la concentración de potasio en la sangre
es otro potente estímulo para la secreción de aldosterona. Además, este efecto se ve potenciado por la
presencia de ANG II.
• Hormona Adrenocorticotropa (ACTH): Aunque la ACTH principalmente estimula la síntesis de
cortisol, también tiene un efecto, aunque menos potente, en la secreción de aldosterona.
Secreción de aldosterona por la acción de la Angiotensina II (Sistema Renina- Angiotensina-
Aldosterona)
1. Hipoperfusión Renal. Cuando hay una disminución en la perfusión sanguínea renal, las células
yuxtaglomerulares, ubicadas en el aparato yuxtaglomerular de las nefronas, detectan esta
disminución y secretan la enzima proteica llamada renina.
2. Secreción de Renina. La renina se libera en la sangre y tiene una vida media de aproximadamente 30
a 60 minutos.
3. Acción de la Renina sobre el Angiotensinógeno. La renina actúa sobre el angiotensinógeno, una
globulina sintetizada en el hígado, para convertirla en angiotensina I. El angiotensinógeno es una
proteína precursora que se encuentra en la circulación sanguínea.
4. Conversión de Angiotensina I a Angiotensina II. La angiotensina I, que es biológicamente inactiva,
se convierte en angiotensina II principalmente a nivel pulmonar gracias a la enzima convertidora de
angiotensina (ECA). Esta enzima es especialmente abundante en los pulmones.
5. Funciones de la Angiotensina II. Tiene varias funciones:
a. vasoconstricción, que aumenta la resistencia vascular y, por lo tanto, la presión arterial.
b. Estimula la liberación de aldosterona de la corteza suprarrenal, lo que aumenta la reabsorción
de sodio y agua en los riñones, contribuyendo así a la regulación del volumen sanguíneo y la
presión arterial.
c. También estimula la liberación de vasopresina (hormona antidiurética), que aumenta la
reabsorción de agua en los riñones.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
La acción de la angiotensina II (ANG II) en la célula glomerular para estimular la síntesis y secreción de
aldosterona implica una cascada de eventos intracelulares. Aquí se detalla el proceso:
6. Unión a Receptor AT1. La ANG II se une al receptor de angiotensina II tipo 1 (AT1) en la membrana
plasmática de la célula glomerular.
7. Activación de la Gαq y Fosfolipasa C (PLC). A través de la mediación de la proteína Gαq, el receptor
AT1 activa la fosfolipasa C (PLC).
8. Formación de Diacilglicerol (DAG) e Inositol 1,4,5-Trifosfato (IP3). La activación de la PLC
desencadena la formación de dos segundos mensajeros: el DAG y el IP3.
a. El DAG activa la proteína quinasa C (PKC), una enzima importante en la transducción de
señales intracelulares.
b. El IP3 provoca la liberación de calcio (Ca2+) desde los depósitos intracelulares, como el
retículo endoplásmico.
9. El aumento de la concentración de Ca2+ en el citoplasma activa diversas enzimas dependientes de
Ca2+, como la PKC y las proteínas quinasas dependientes de Ca2+-calmodulina.
10. Despolarización de la Membrana Plasmática. Los cambios intracelulares mencionados resultan en la
despolarización de la membrana plasmática de la célula glomerular.
11. Apertura de Canales de Ca2+ Activados por Voltaje. La despolarización permite la apertura de
canales de Ca2+ activados por voltaje, generando una entrada prolongada de Ca2+ desde el espacio
extracelular.
12. Estimulación de la Síntesis y Secreción de Aldosterona. El aumento de la concentración intracelular
de Ca2+ facilita la síntesis de pregnenolona al aumentar la actividad de la enzima limitante de
velocidad, la esteroidogénesis aguda de la zona glomerular (SCC). Además, estimula la aldosterona
sintasa, favoreciendo la conversión de corticosterona en aldosterona.
Secreción de la aldosterona por incremento de potasio.
1. Canales de Potasio (K+). Las células glomerulares tienen varios canales de potasio que son
responsables de mantener el potencial de reposo normal de la membrana.
2. Despolarización de la Membrana. Un aumento en [K+]e induce la despolarización de la membrana
plasmática de la célula glomerular.
3. Apertura de Canales de Ca2+ Voltaje-Dependientes. La despolarización resultante abre los canales de
calcio (Ca2+) voltaje-dependientes en la membrana celular.
4. Entrada de Ca2+. La apertura de los canales de Ca2+ permite la entrada de iones de calcio desde el
espacio extracelular hacia el citoplasma.
5. Estimulación de la Producción de Pregnenolona y Conversión a Aldosterona. El aumento de la
concentración intracelular de Ca2+ estimula la producción de pregnenolona a partir del colesterol y
la conversión de corticosterona en aldosterona.
Es importante destacar que, a diferencia de la acción de la angiotensina II (ANG II), el aumento de [K+]e no
requiere la activación de la fosfolipasa C (PLC) ni la liberación de Ca2+ desde los depósitos intracelulares.
Ambos, ANG II y el aumento de [K+]e, comparten el mecanismo de aumentar la concentración intracelular
de Ca2+, lo que sugiere una posible acción sinérgica entre estos estímulos en las células glomerulares.
Secreción de aldosterona por incremento de ACTH.
La hormona adrenocorticotropa (ACTH) estimula la secreción de aldosterona a través de una vía distinta en
comparación con la angiotensina II (ANG II) y el aumento de potasio extracelular ([K+]e). Aunque el efecto
de la ACTH es mínimo en comparación con estos estímulos, desencadena eventos específicos en las células
glomerulares:
1. Receptor MC2R y Adenilato Ciclasa. En las células glomerulares, el receptor MC2R se acopla a la
adenilato ciclasa mediante una proteína G. Cuando llega la ACTH se une con este receptor acoplado
a proteína G la cual activa la AC produciendo un aumento de AMPc
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
2. Aumento de AMPc (Monofosfato de Adenosina Cíclico). La elevación de la concentración
intracelular de AMPc activa la proteína quinasa A (PKA).
3. Activación de la PKA. La PKA, una vez activada, fosforila numerosas proteínas en el citosol de la
célula glomerular.
4. Entrada de Ca2+. A través de eventos aún por definir, estos cambios inducen la entrada de calcio
(Ca2+) a través de la membrana plasmática.
5. Estimulación de la Síntesis y Secreción de Aldosterona. El aumento de la concentración intracelular
de Ca2+ estimula la síntesis y secreción de aldosterona por las células glomerulares.
Además de este mecanismo, la ACTH también incrementa la actividad mineralocorticoide a través de otro
proceso. Estimula las células fasciculares para que secreten cortisol, corticosterona y DOC, todos los cuales
tienen una leve actividad mineralocorticoide. Se postula que la hipertensión observada en individuos con un
exceso de secreción de ACTH se debe a la síntesis excesiva de estos mineralocorticoides débiles.
Tratamiento del aumento de aldosterona mediante el eje RAA.
En el tratamiento farmacológico del aumento de aldosterona podemos destacar:
• Antagonistas de la Aldosterona (Espironolactona y Eplerenona): Estos fármacos, que son diuréticos,
bloquean la acción de la aldosterona a nivel del receptor mineralocorticoide (MR). Aunque son
diuréticos débiles, resultan especialmente útiles en pacientes con hipertensión resistente al
tratamiento. En casos de hipertensión resistente o insuficiencia cardíaca congestiva, se suele
combinar un antagonista de la aldosterona con un diurético tiazídico para evitar la pérdida excesiva
de potasio.
• Inhibidores de la ECA (Enzima Convertidora de Angiotensina): Estos fármacos bloquean la enzima
que convierte la angiotensina I en angiotensina II. Los inhibidores de la ECA son considerados
seguros y bien tolerados. Además de tratar la hipertensión, estos medicamentos mejoran la calidad de
vida y la supervivencia en pacientes con insuficiencia cardíaca.
• Antagonistas de los Receptores de Angiotensina (ARA): Estos fármacos bloquean específicamente
los receptores AT1 a los que se une la angiotensina II. Ofrecen una alternativa válida para aquellos
pacientes que no toleran los inhibidores de la ECA. Al igual que los inhibidores de la ECA, los ARA
son efectivos en el tratamiento de la hipertensión y pueden tener beneficios adicionales en
condiciones como la insuficiencia cardíaca.
• Inhibidores Directos de la Renina: Estos medicamentos actúan en la fase inicial del sistema renina-
angiotensina al inhibir directamente la renina. Al bloquear la formación de angiotensina I, reducen la
producción de angiotensina II y, por ende, la liberación de aldosterona. Esta clase de fármacos es una
adición más reciente al arsenal terapéutico para la hipertensión.
8.3.- Andrógenos
• Las células de la capa reticular son las encargadas de fabricar andrógenos, que son hormonas
importantes. Utilizan sustancias llamadas 17α-hidroxipregnenolona y 17α-hidroxiprogesterona para
producir andrógenos específicos como la dehidroepiandrosterona (DHEA) y la androstenediona.
• Enzima 17,20-desmolasa. Esta enzima convierte esas sustancias que mencionamos antes en los
andrógenos que necesitamos.
• Potencia de los Andrógenos Suprarrenales. Los andrógenos que fabricamos aquí no son tan fuertes
como la testosterona o la dihidrotestosterona, pero son importantes. Otros tejidos, como el hígado y
los riñones, pueden convertir uno de estos andrógenos (llamado androstenediona) en testosterona.
• Contribución de las Suprarrenales. Las glándulas suprarrenales, en especial en las mujeres, aportan
significativamente a los andrógenos que circulan en el cuerpo. Esto es especialmente relevante en un
período antes de la pubertad, llamado adrenarquia, que precede a la activación gonadal durante la
pubertad. Este aumento en la producción de andrógenos estimula el crecimiento del vello púbico y
axilar.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
8.4.- Médula suprarrenal.
Las células de la médula, denominadas células cromafines debido a que las catecolaminas que contienen
muestran una gran avidez por la tinción con sales de cromo; sintetizan y secretan adrenalina y, en menor
medida, noradrenalina. La noradrenalina es el neurotransmisor de la división simpática del sistema nervioso
autónomo. Tanto la noradrenalina como la adrenalina producidas en la médula suprarrenal acceden a la
circulación y actúan sobre los tejidos distales, igual que cualquier hormona.
Las fibras simpáticas posganglionares de los nervios esplácnicos, que liberan acetilcolina (ACh), son los
principales reguladores de la secreción hormonal por la médula suprarrenal. Destacar que estas células
presentan una irrigación peculiar que expone la médula suprarrenal a las mayores concentraciones de
glucocorticoides y mineralocorticoides en comparación con otros tejidos somáticos.
Síntesis de catecolaminas.
• Tirosina Hidroxilasa. Esta enzima juega un papel clave al
convertir el aminoácido tirosina en l-dopa. Se encuentra en diferentes
lugares importantes, como las células de la médula suprarrenal, los
terminales nerviosos simpáticos y ciertas células del sistema nervioso
central. La síntesis global de estas hormonas comienza aquí, y la actividad
de esta enzima determina la velocidad del proceso.
• Aminoácido Descarboxilasa Citosólica. Después, otra enzima,
conocida como aminoácido descarboxilasa citosólica, transforma la l-dopa
en dopamina. Este paso ocurre en varios tejidos, incluida la médula
suprarrenal.
• Un intercambiador de catecolaminas-H + traslada la dopamina al
interior de unas vesículas membranosas de núcleo denso llamadas gránulos
cromafines.
• Dopamina β-Hidroxilasa. Dentro de estos gránulos, la dopamina
es convertida en noradrenalina por la enzima dopamina β-hidroxilasa. Esto
ocurre específicamente en la médula suprarrenal y en los nervios simpáticos.
• La noradrenalina formada en los gránulos secretores sale hacia el
citosol.
• Feniletanolamina N-Metiltransferasa (PNMT). Una enzima
llamada feniletanolamina N-metiltransferasa (PNMT), presente en el citosol
de las células cromafines suprarrenales, juega un papel fundamental en la
conversión de noradrenalina a adrenalina. Esta enzima solo existe en una
cantidad importante en el citosol de las células cromafines suprarrenales.
• Absorción y Almacenamiento en Gránulos. La adrenalina recién
formada se absorbe en los gránulos secretorios de la médula suprarrenal. Se piensa que un
intercambiador de catecolaminas-H+ facilita este proceso. Estos gránulos mantienen una reserva de
adrenalina y noradrenalina, listos para ser liberados en respuesta a situaciones de estrés.
Degradación catecolaminas.
1. Acciones Breves. Las catecolaminas, como la
adrenalina, tienen acciones biológicas muy breves,
alrededor de 10 segundos.
2. Primera Etapa de Degradación
a. Enzima: Catecol-O-metiltransferasa (COMT)
b. Localización: Células endoteliales, corazón,
hígado, riñones.
c. Acción:
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
i. Adrenalina se convierte en metanefrina.
ii. Noradrenalina se convierte en normetanefrina.
3. Segunda Etapa de Degradación.
a. Enzima: Monoaminooxidasa
b. Acción: Metanefrina y normetanefrina se convierten en ácido vanililmandélico (AVM).
4. El hígado y el intestino se encargan de unir estos productos de degradación a sulfato o glucuronato,
formando derivados. Estos derivados son excretados por el riñón a través de la orina.
Activación de la síntesis de catecolaminas.
1. Estimulación por ACTH. ACTH estimula la síntesis de l-dopa y noradrenalina.
2. Regulación Positiva por Cortisol. El cortisol, transportado desde la corteza suprarrenal, regula
positivamente la feniletanolamina N-metiltransferasa (PNMT) en células cromafines. Resulta en una
sinergia entre los ejes CRH-ACTH-Cortisol y simpático-adrenalina, prolongando la respuesta al
factor estresante.
En los gránulos secretorios de la médula suprarrenal, las catecolaminas se almacenan junto con ATP y Ca2+,
unidas a proteínas granulares llamadas cromograninas. La liberación de catecolaminas está controlada por el
sistema nervioso central (SNC). La liberación se desencadena por la acción de la acetilcolina (ACh) de las
neuronas preganglionares en receptores nicotínicos, desencadenando una serie de eventos que culminan en
la liberación de adrenalina por exocitosis. Adicionalmente, las cromograninas, especialmente Cromogranina
B, contribuyen a la densidad del núcleo denso de las vesículas, y la liberación de Cromogranina A se utiliza
como indicador de la actividad de la médula suprarrenal.
En la circulación, las catecolaminas, liberadas en respuesta al estrés, se disocian del complejo de unión para
actuar libremente sobre los tejidos diana, desencadenando respuestas fisiológicas necesarias para enfrentar
situaciones de estrés.
Vías de acción intracelular de las catecolaminas
La adrenalina y la noradrenalina ejercen sus efectos al unirse a receptores adrenérgicos, todos los cuales
pertenecen a la familia de receptores acoplados a proteínas G (GPCR). Estos receptores, a su vez, pueden
reconocer tanto la adrenalina como la noradrenalina, aunque con afinidades diferentes. La versatilidad de
estas sustancias resulta en efectos tanto excitadores como inhibidores, dependiendo del tejido específico.
• Receptores α y β:
Se distinguen receptores α-adrenérgicos, asociados principalmente con la estimulación, y receptores β-
adrenérgicos, asociados principalmente con la inhibición.
• Subtipos de Receptores:
o Receptores β:
▪ Se conocen al menos tres tipos de receptores β (β1, β2, β3).
▪ Acoplados a proteínas G heterotriméricas estimuladoras (Gαs) que activan la
adenilatociclasa, aumentando los niveles de AMPc, el principal mediador intracelular
de la activación β.
o Receptores α:
▪ Se reconocen al menos dos tipos de receptores α (α1, α2).
▪ Receptores α2 se asocian a proteínas G inhibidoras (Gαi) que disminuyen los niveles
de AMPc en los tejidos diana.
▪ Receptores α1 se acoplan a proteínas G heterotriméricas (Gαq) que activan la
fosfolipasa C (PLC), aumentando la concentración intracelular de IP3 y Ca2+.
La amplia variedad de receptores adrenérgicos y sus subtipos permite respuestas específicas en diferentes
tejidos, desde la estimulación hasta la inhibición. La conexión con distintas proteínas G juega un papel
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
crucial en la transducción de señales y determina la naturaleza de la respuesta celular a la estimulación
adrenérgica.
Acciones de las catecolaminas en el ejercicio.
Algunas de las acciones atribuidas exclusivamente a la liberación suprarrenal de catecolaminas que integran
distintas funciones corporales como parte de la respuesta al estrés durante el ejercicio son:
• Aumento del Flujo Sanguíneo Muscular. La adrenalina contribuye al incremento del flujo sanguíneo
hacia los músculos durante el ejercicio, optimizando el suministro de oxígeno y nutrientes
necesarios.
• Relajación del Músculo Liso Bronquial. La relajación mediada por la adrenalina en el músculo liso
bronquial facilita una mayor ventilación, mejorando la capacidad respiratoria durante el ejercicio.
• Degradación de Glucógeno Muscular. Activación del adrenorreceptor β inicia la degradación de
glucógeno en los músculos, proporcionando un suministro inmediato de combustible para la
contracción muscular.
• Lipólisis en Tejido Adiposo. La adrenalina activa la lipólisis en el tejido adiposo, liberando ácidos
grasos para servir como fuente adicional de energía durante el ejercicio.
• Activación de Glucogenólisis en el Hígado. En el hígado, la adrenalina estimula la glucogenólisis,
asegurando un suministro constante de glucosa a la sangre para satisfacer las demandas energéticas
del cuerpo.
• Regulación de la Insulina durante el Ejercicio. La adrenalina, a través del adrenorreceptor β,
inicialmente estimula la secreción de insulina, pero la inervación autónoma local bloquea este efecto
durante el ejercicio, favoreciendo la glucogenólisis y manteniendo la glucemia.
Feocromocitoma
Un feocromocitoma es un tumor relativamente poco frecuente producido por la hiperplasia o, rara vez,
neoplasia de tejido medular suprarrenal o de tejido cromafín extrasuprarrenal que no involucionó tras el
parto. Estos tumores, que pueden ser benignos o malignos, fabrican catecolaminas al igual que la médula
normal, pero sin regulación de ningún tipo. Los pacientes con feocromocitomas suelen mostrar una gran
cantidad de síntomas, como es de esperar en un sistema hormonal de tan amplio alcance. Entre los datos
clínicos destacan: hipertensión paroxística (súbita), taquicardia, cefalea, episodios de sudoración, ansiedad,
temblores e intolerancia a la glucosa. La clave para diagnosticar este trastorno es una anamnesis detallada,
los datos de la exploración física que indiquen un exceso de tono adrenérgico y la determinación analítica de
unas cantidades elevadas de catecolaminas y sus metabolitos tanto en la sangre como en la orina. Una vez
que el análisis químico de los metabolitos confirma la presencia de un feocromocitoma, en ocasiones se
puede localizar el tumor en una de las glándulas suprarrenales y extirparlo. En raras ocasiones ambas
glándulas se encuentran afectadas y se requiere una adrenalectomía bilateral. Posteriormente, estos pacientes
deberán ser tratados con glucocorticoides y mineralocorticoides sustitutivos. No se administra tratamiento
alguno para reemplazar la función medular suprarrenal. No se sabe bien si estos sujetos reaccionan menos a
los estímulos externos que desencadenan la respuesta de lucha o huida.
9.- PÁNCREAS ENDOCRINO.
El páncreas es un órgano dual con funciones tanto exocrinas como endocrinas. Mientras que las glándulas
exocrinas liberan enzimas digestivas y bicarbonato hacia la luz intestinal, las glándulas endocrinas forman
los llamados islotes de Langerhans. Características de los Islotes de Langerhans:
• Distribución y Proporción:
o Constituyen solo el 1-2% de la masa total del páncreas.
o Diseminados por todo el páncreas, con un páncreas humano normal que contiene entre
500,000 y varios millones de islotes.
o Los islotes pueden ser ovales o esféricos y miden entre 50 y 300 µm de diámetro.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• Tipos de Células Secretoras:
o Células β: Son el tipo más abundante y se encuentran especialmente en el centro del islote.
Secretan insulina, proinsulina, péptido C y amilina (IAPP).
o Células α: Secretan principalmente glucagón.
o Células δ: Secretan somatostatina.
o Células F: También conocidas como células productoras de polipéptido pancreático, secretan
polipéptido pancreático.
• Productos Secretados:
o Insulina: Facilita la absorción de glucosa por las células.
o Proinsulina: Se convierte en insulina y péptido C durante la secreción.
o Péptido C: Se utiliza como marcador para evaluar la producción de insulina.
o Amilina (IAPP): Recientemente descubierta, su función precisa aún se está investigando.
o Glucagón: Aumenta los niveles de glucosa en sangre.
o Somatostatina: Inhibe la liberación de insulina y glucagón, regulando la función pancreática.
Comunicación entre Células de los Islotes de Langerhans.
• Comunicación Humoral:
o Irrigación del Islote: La irrigación sanguínea transcurre del centro hacia la periferia del islote,
llevando consigo glucosa y otros secretagogos.
o Distribución Celular: En algunos casos, las células β son más abundantes en el centro del
islote, mientras que las células α y δ predominan en la periferia.
o Influencia Mutua: Las células de un islote pueden afectar la secreción de otras células a
medida que la sangre lleva los productos hormonales secretados por cada tipo celular.
o Ejemplos de Interacción. Glucagón es un potente estimulador de la liberación de insulina. La
insulina inhibe moderadamente la liberación de glucagón.Somatostatina inhibe fuertemente la
secreción tanto de insulina como de glucagón, así como de otras hormonas no producidas en
los islotes.
• Comunicación de Célula a Célula:
o Uniones Celulares. Las células de los islotes están conectadas por uniones estrechas y
uniones en hendidura.
o Importancia: Las comunicaciones a través de estas uniones pueden ser cruciales para regular
la secreción de insulina y glucagón.
• Comunicación Neural:
o Regulación Autónoma: La secreción de los islotes está regulada por las divisiones simpática y
parasimpática del sistema nervioso autónomo (SNA).
o Estimulación Colinérgica: Incrementa la secreción de insulina.
o Estimulación Adrenérgica: Puede tener un efecto estimulador o inhibidor, dependiendo de si
predomina la estimulación β-adrenérgica (estimuladora) o α-adrenérgica (inhibidora).
9.1.- Insulina.
Síntesis de insulina.
La insulina es una hormona crucial en la regulación de los niveles de glucosa en sangre. Aquí está una
explicación detallada del proceso de síntesis de insulina:
• Gen de la Insulina. La insulina es codificada por un solo gen localizado en el brazo corto del
cromosoma 11.
• Estimulación por la Glucosa. La síntesis y secreción de insulina son estimuladas por la exposición de
los islotes pancreáticos a la glucosa. La glucosa debe metabolizarse para que se produzca esta
estimulación, aunque los detalles precisos de este proceso aún no se comprenden completamente.
• Síntesis de Insulina.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
o La transcripción del gen de la insulina da lugar al ARN
mensajero (ARNm) completo que codifica la preproinsulina.
o La preproinsulina contiene un péptido señal en su extremo 5',
seguido de los dominios peptídicos B, C y A.
o Durante la síntesis, en el retículo endoplásmico rugoso, el
péptido señal de aproximadamente 24 aminoácidos se escinde
de la preproinsulina, generando la proinsulina.
• Procesamiento en el Aparato de Golgi:
o En el aparato de Golgi, la proinsulina es empaquetada en
gránulos secretorios.
o En esta etapa, las proteasas comienzan a escindir la proinsulina
en dos lugares, eliminando el péptido C de 31 aminoácidos.
o La molécula resultante de insulina tiene dos cadenas peptídicas,
A de 21 aminoácidos y B de 30 aminoácidos, unidas por dos
puentes disulfuro.
• Empaquetado y Secreción:
o En el gránulo secretor, la insulina se asocia con zinc.
o La vesícula secretora contiene insulina, proinsulina y péptido C.
Estos se liberan en la sangre portal cuando las células beta son
estimuladas por la glucosa.
Secreción de insulina, proinsulina y péptido C
El péptido C, aunque carece de función biológica conocida, se presenta en una proporción equitativa
(proporción molar1:1) con la insulina al ser secretado, convirtiéndolo en un indicador valioso de la actividad
insulínica. La proinsulina, aunque menos potente que la insulina, se secreta en pequeñas cantidades
(alrededor del 5% de la insulina). La medición directa de la concentración de insulina no refleja con
precisión la cantidad liberada, ya que aproximadamente el 60% de la insulina secretada en la circulación
portal es eliminada por el hígado en una primera etapa. En contraste, el péptido C no es captado por el
hígado y su medición proporciona una estimación más precisa de la insulina liberada, ya que se elimina en la
orina. La cantidad de péptido C excretado en 24 horas sirve como una aproximación de la actividad
insulínica en ese período.
La glucosa es el principal regulador de la secreción de insulina.
En individuos sanos, la concentración de glucosa en plasma se mantiene en rangos muy estrechos,
normalmente entre 4 y 5 mM después de un ayuno nocturno. Tras las comidas, la concentración puede
aumentar, pero incluso con comidas abundantes, generalmente no supera los 10 mM. Pequeños aumentos en
la glucosa desencadenan una fuerte secreción de insulina y péptido C, elevando la concentración de insulina,
como se observa en una prueba de tolerancia oral a la glucosa (PTOG). Por el contrario, una disminución del
20% en la glucosa reduce significativamente la concentración de insulina. El cambio en la glucosa
plasmática ya sea después de comer o durante el ayuno, es el principal factor que regula la secreción de
insulina. Cuando se administra una carga de glucosa intravenosa (0,5 g/kg de peso corporal), la
concentración de glucosa en plasma aumenta más rápidamente que cuando se administra por vía oral. Este
rápido aumento lleva a dos fases distintas en la secreción de insulina.
• Fase Aguda (Primera Fase):
o Estímulo: Incremento rápido de glucosa (ej. comida).
o Duración: 2-5 minutos.
o Fuente de Insulina:
o Vesículas preformadas cercanas a la membrana plasmática.
o Participación del Péptido C:
o Secreción proporcional a la insulina.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• Fase Crónica (Segunda Fase):
o Estímulo: Persistencia del aumento de glucosa.
o Duración: Hasta normalización de niveles de glucemia.
o Fuente de Insulina:
o Vesículas preformadas y, en parte, de nueva síntesis.
o Participación del Péptido C: Contribuye, pero en menor medida que en la fase aguda.
• Nota:
o En condiciones normales, ambas fases contribuyen a la respuesta total de insulina.
o En la diabetes tipo 1 y tipo 2, la pérdida de la primera fase es un primer indicio detectable.
El metabolismo de la glucosa por la célula β estimula la secreción de insulina.
Las células β del páncreas responden a múltiples estímulos para regular su liberación. La glucosa es el
principal inductor, desencadenando la secreción de insulina durante su metabolización. Otras sustancias,
como galactosa y manosa, también pueden estimular esta respuesta. Sin embargo, algunas hexosas no
metabolizables, como la 3-O-metilglucosa o la 2-desoxiglucosa, no generan la liberación de insulina.
Además de la glucosa, aminoácidos como la arginina y la leucina, junto con pequeños cetoácidos, pueden
tener un efecto estimulante en la secreción de insulina, aunque su influencia es menor en comparación con la
glucosa. La fructosa, una cetohexosa, también puede inducir moderadamente la secreción de insulina. Estos
diversos estímulos permiten una respuesta adaptable para mantener la homeostasis de glucosa y otros
sustratos energéticos en la sangre.
La liberación de insulina en respuesta a la glucosa sigue un proceso en siete etapas dentro de las células β
del páncreas:
1. La glucosa entra en la célula β a través del transportador de glucosa GLUT2 mediante difusión
facilitada.
2. En presencia de glucocinasa, la glucosa experimenta glucólisis y oxidación por el ciclo del ácido
cítrico, generando ATP y aumentando las proporciones de [ATP]i/[ADP]i, [NADH]i/[NAD+]i y
[NADPH]i/[NADP+]i.
3. El aumento en estas proporciones cierra los canales KATP.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Los canales KATP (canales de potasio
sensibles al ATP) son canales iónicos
presentes en las membranas de varias células,
incluidas las células β del páncreas. Cuando la
concentración intracelular de ATP es alta,
como ocurre en situaciones de alta
disponibilidad de glucosa y en la presencia de
otros nutrientes, los canales KATP se cierran,
disminuyendo la conductancia de K+
4. La reducción de la conductancia de K+
despolariza la célula β, haciendo que el
potencial de membrana sea menos
negativo.
5. La despolarización activa los canales
de Ca2+ dependientes de voltaje.
6. La mayor permeabilidad al Ca2+
provoca una mayor entrada de Ca2+,
elevando el Ca2+ libre intracelular y
estimulando la liberación de Ca2+
inducida por Ca2+.
7. El aumento del Ca2+ intracelular,
posiblemente mediante la activación
de la cascada de fosforilación Ca2+-
calmodulina, finalmente induce la liberación de insulina.
Además de la vía descrita anteriormente, la secreción de insulina puede ser modulada por otros secretagogos
a través de diferentes vías de señalización celular. Algunos de estos secretagogos actúan a través de la vía de
la fosfolipasa C o de la vía de la adenilato ciclasa, en lugar de depender exclusivamente de la vía de la
glucosa y el calcio.
Por ejemplo, el glucagón, que estimula la liberación de insulina, puede activar la adenilato ciclasa,
aumentando así los niveles de AMPc y activando la proteína cinasa A (PKA), sorteando parcial o
completamente la vía de la glucosa y el calcio. En contraste, la somatostatina, que inhibe la liberación de
insulina, puede actuar mediante la inhibición de la adenilato ciclasa.
La secreción de insulina es modulada por factores neurales y humorales.
• Inervación:
o Simpática. Estimulación β-adrenérgica aumenta insulina. En situaciones de estrés, la
activación del sistema simpático prepara al cuerpo para afrontar demandas energéticas
elevadas.
o Parasimpática. Estimulación α-adrenérgica inhibe insulina. Durante períodos de reposo o
digestión, la acción del sistema parasimpático reduce la liberación de insulina.
• Ejercicio:
o Inhibición de la liberación de insulina por la activación de los receptores α-adrenérgicos
durante el ejercicio evita hipoglucemia. Lo que se conoce como inhibición α-adrenérgica
o Permite al músculo utilizar glucosa eficientemente incluso con bajos niveles de insulina.
o Evita la captación excesiva de glucosa que podría llevar a una peligrosa reducción de azúcar
en sangre.
o Al inhibir la insulina, se reduce la captación de glucosa por tejidos como el tejido adiposo,
permitiendo que la glucosa circule en mayor cantidad en el torrente sanguíneo y esté
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
disponible para los músculos activos.
Al hacer ejercicio, el tejido muscular utiliza glucosa aunque la concentración plasmática de insulina sea
baja. Si los niveles de insulina aumentan, la utilización de glucosa por parte del músculo aumentaría aún
más, provocando hipoglucemia. Además, un aumento en la concentración de insulina inhibiría la lipolisis y
la liberación de ácidos grasos desde los adipocitos, reduciendo así la disponibilidad de ácidos grasos, que
el músculo podría utilizar como combustible alternativo a la glucosa. Por último, una elevación de los
niveles de insulina reduciría la producción de glucosa por el hígado. De este modo, la supresión de la
secreción de insulina durante el ejercicio evita que el músculo capte un exceso de glucosa, que, si superase
la capacidad hepática para producir glucosa, provocaría una hipoglucemia grave con afectación del
cerebro
• Ingesta de Alimentos:
o Fase Cefálica:
▪ Antes de la ingesta, liberación de insulina estimulada por nervio vago.
▪ El nervio vago activa la liberación de insulina en anticipación a la llegada de
alimentos, preparando al cuerpo para el aumento inminente de glucosa.
o Ingesta de Comida:
▪ Acetilcolina liberada por nervio vago aumenta respuesta insulínica a glucosa. La
acetilcolina actúa sobre receptores muscarínicos. La activación de receptores
muscarínicos estimula la producción de segundos mensajeros, como el inositol
trifosfato (IP3) y el diacilglicerol (DAG). Estos segundos mensajeros participan en la
liberación de calcio intracelular. El aumento del calcio intracelular es crucial para la
secreción de insulina. El calcio activa maquinaria celular que conduce a la exocitosis
de vesículas secretoras que contienen insulina. La liberación de insulina en respuesta
a la estimulación parasimpática (vía acetilcolina) contribuye a la regulación de la
homeostasis glucémica.
▪ Facilita la captación de glucosa, permitiendo que las células beta respondan
eficazmente al aumento repentino de niveles de azúcar en sangre.
• Incretinas (Péptidos Intestinales):
o Colecistocinina, GLP-1, GIP. Liberados por células intestinales en respuesta a la ingesta de
alimentos.
o GLP-1 (Péptido Similar al Glucagón Tipo 1):
▪ Importante incretina que mejora la respuesta de las células beta a la glucosa.
▪ Vida media corta lo que limita su uso terapéutico.
▪ En la saliva del monstruo de Gila, se encontró un péptido parecido al GLP-1,
conocido como exendina-4. Para tratar la diabetes, se ha creado una versión sintética
de larga duración llamada exenatida. Este compuesto se administra mediante
inyección subcutánea, ya que, al ser un péptido, no puede tomarse por vía oral.
Además, existen medicamentos de administración oral llamados gliptinas, que inhiben
la enzima dipeptidil-peptidasa 4, encargada de degradar el GLP-1 natural. Estas
gliptinas se utilizan en el tratamiento de la diabetes tipo 2.
Efecto Incretina.
La administración intravenosa no activa completamente el efecto incretina, ya que este fenómeno está
específicamente asociado con la estimulación de las células beta del páncreas en respuesta a los nutrientes
que ingresan al sistema digestivo.
Las incretinas, como el GLP-1 (péptido similar al glucagón tipo 1) y el GIP (péptido inhibidor gástrico), son
liberadas en el tracto gastrointestinal en respuesta a la ingestión de alimentos. Estos péptidos entran en
circulación y afectan positivamente la secreción de insulina, entre otras funciones.
El receptor de insulina es un receptor tirosina-cinasa
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Una vez secretada en la sangre portal, la insulina inicialmente llega al hígado, donde se retiene más de la
mitad y es retirada de la circulación. La insulina que escapa del hígado está disponible para desencadenar
procesos sensibles a la insulina en otros tejidos. En cada tejido diana, la primera acción de la insulina es
unirse a un receptor específico de tirosina-cinasa en la membrana plasmática.
El receptor de insulina es un heterotetrámero compuesto por dos cadenas α y dos cadenas β. Estas cadenas
están unidas por puentes disulfuro y tienen similitud estructural con el receptor de IGF-1 (receptor del factor
de crecimiento). El receptor de insulina tiene una región de tirosina-cinasa crucial para su función. La
insulina se une a las cadenas α del receptor, lo que activa la tirosina-cinasa en las cadenas β.
Así pues, el receptor de insulina puede fosforilarse a sí mismo y a otros sustratos intracelulares, como las
proteínas IRS (sustratos del receptor de insulina) y SHC (proteínas homólogas con el dominio C-terminal de
Src). La fosforilación de tirosinas en estas proteínas, especialmente en IRS-1, activa múltiples vías de
señalización simultáneamente. Estas vías son fundamentales para transmitir la señal de la insulina a través
de la membrana plasmática de los tejidos diana (no son muy importantes pq no vienen en la diapositiva).
1. Vía PI3K (Fosfatidilinositol 3-cinasa):
a. Fosforilación de IRS: La insulina se une a su receptor, desencadenando la fosforilación de
IRS (sustrato del receptor de insulina).
b. Activación de PI3K: IRS fosforilado activa PI3K, que convierte el fosfatidilinositol-4,5-
bisfosfato (PIP2) en fosfatidilinositol-3,4,5-trisfosfato (PIP3).
c. PDK y Activación de PKB: PIP3 activa PDK (cinasa dependiente de fosfatidilinositol), que a
su vez activa PKB (proteína-cinasa B) mediante fosforilación.
d. Impacto:
i. Translocación de GLUT4: PKB facilita la translocación de transportadores de
glucosa GLUT4 a la membrana celular, aumentando la captación de glucosa.
ii. Inhibición de GSK-3: PKB fosforila e inactiva GSK-3, disminuyendo la inhibición
de la glucógeno-sintasa y favoreciendo la síntesis de glucógeno.
iii. mTOR y Traducción de Proteínas: Activación de mTOR, influyendo en la
traducción de ARNm y la síntesis proteica.
2. Vía de Proteínas Ras y MAPK:
a. Fosforilación de SHC: El receptor de insulina fosforila SHC.
b. Activación de GRB2: La proteína GRB2 se activa, estimulando la vía Ras.
c. Activación de SOS y Ras: SHC y GRB2 activan SOS, que activa Ras.
d. Cascada MAPK (Ras-MEK-MAPK): Activación de MEK y MAPK (cinasas) que afectan la
expresión génica y la actividad de otras cinasas.
e. Impacto:
i. Expresión Génica: La vía Ras-MAPK regula la expresión de genes involucrados en
procesos celulares, como la proliferación y diferenciación.
ii. p90-S6 Cinasa y Glucógeno-Sintasa: La cascada activa p90-S6 cinasa y afecta la
actividad de glucógeno-sintasa mediante la fosforilación de PP1.
3. Vía con Proteínas SH2:
a. Unión a Secuencias Fosfotirosina: Proteínas con dominios SH2 se unen a secuencias
fosfotirosina específicas en el receptor de insulina o en IRS.
b. Respuestas Celulares Variadas: Las proteínas SH2 regulan diversas rutas celulares, incluido el
metabolismo lipídico y otras respuestas intracelulares.
Efectos de la insulina.
Tras la ingesta de alimento aumenta la concentración plasmática de insulina, estimulada por la glucosa y por
la activación, neural y mediante las incretinas, de las células β; las tres dianas principales de la acción de la
insulina son el hígado, el músculo y el tejido adiposo:
Hígado:
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
1. Captación y Almacenamiento de Glucosa:
• La insulina estimula la captación de glucosa por el hígado mediante el transportador GLUT2.
• Incrementa la glucogénesis, promoviendo la síntesis de glucógeno a partir de glucosa.
• Inhibe la glucogenólisis, reduciendo la descomposición de glucógeno en glucosa.
• Inhibe la gluconeogénesis, disminuyendo la formación de glucosa a partir de precursores no
glucídicos.
2. Regulación del Metabolismo Lipídico:
• Favorece la lipogénesis, estimulando la síntesis de ácidos grasos y la formación de
triglicéridos.
• Inhibe la oxidación de ácidos grasos al reducir la actividad de la carnitina-acetiltransferasa I,
bloqueando así la entrada de ácidos grasos en las mitocondrias.
• Estimula la síntesis de apolipoproteínas para la formación de VLDL, participando en el
transporte de triglicéridos desde el hígado.
3. Influencia en el Metabolismo Proteico:
• Estimula la síntesis proteica.
• Inhibe la degradación de proteínas en el hígado.
Músculo:
1. Captación y Almacenamiento de Glucosa:
• La insulina induce la movilización de transportadores GLUT4 hacia la membrana plasmática,
aumentando la captación de glucosa.
• Estimula la conversión de glucosa en glucógeno a través de la activación de la hexocinasa y
la glucógeno-sintasa.
2. Regulación del Metabolismo Energético:
• Aumenta la glucólisis y la oxidación de glucosa mediante la activación de la
fosfofructocinasa y la piruvato-deshidrogenasa.
• Estimula la síntesis proteica y reduce la degradación de proteínas, conservando la masa
proteica muscular.
Tejido Adiposo:
1. Captación y Almacenamiento de Glucosa:
• La insulina moviliza GLUT4 hacia la membrana plasmática, facilitando la entrada de glucosa
en el adipocito.
2. Metabolismo de Lípidos:
• Induce la glucólisis, transformando glucosa en metabolitos utilizados para sintetizar
triglicéridos (TG).
• Estimula la síntesis de TG al favorecer la esterificación de ácidos grasos con glicerol.
• Limita la actividad de la lipasa de triglicéridos (ATGL) y la lipasa sensible a hormonas
(HSL), inhibiendo así la descomposición de TG.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
3. Síntesis y Liberación de Triglicéridos:
• Estimula la síntesis de lipoproteína lipasa (LPL), que actúa en la superficie celular,
participando en la conversión de TG de quilomicrones y VLDL en ácidos grasos y glicerol.
• Favorece la captación de ácidos grasos por los adipocitos, promoviendo la formación de
nuevos TG.
9.2.- Glucagón.
Estímulo de Secreción:
• Ingesta de Proteínas: Se considera que la principal señal para la liberación de glucagón es la ingesta
de proteínas. Los aminoácidos liberados durante la digestión de proteínas son importantes
desencadenantes de la secreción de glucagón.
Acciones Principales del Glucagón:
1. Acciones en el Hígado:
• Estimulación de Glucogenólisis: El glucagón promueve la descomposición del glucógeno
en glucosa, aumentando así los niveles de glucosa en la sangre.
1. El glucagón activa la fosforilasa-cinasa mediante fosforilación.
2. La fosforilasa-cinasa activada fosforila la glucógeno-fosforilasa b, convirtiéndola en
la forma activa, glucógeno-fosforilasa a.
3. La fosforilasa a es altamente efectiva en la degradación del glucógeno en glucosa,
aumentando así la glucogenólisis.
• Estimulación de Gluconeogénesis: Facilita la formación de glucosa a partir de precursores
no glucídicos, como aminoácidos y glicerol.
1. El glucagón estimula la gluconeogénesis a través de efectos genómicos.
2. La PKA fosforila el factor de transcripción CREB, que interactúa con elementos de
respuesta al AMPc (CRE), aumentando la expresión de enzimas clave en la
gluconeogénesis (por ejemplo, G6Pasa y PEPCK).
3. CREB fosforilado también estimula la expresión del coactivador-1α del receptor
activado por proliferadores de peroxisomas-γ (PGC-1α), aumentando aún más la
expresión de enzimas gluconeogénicas.
• Cetogénesis: Durante el ayuno, el glucagón favorece la cetogénesis, proporcionando cuerpos
cetónicos como fuente de energía al cerebro.
2. Acciones en el Músculo:
• Glucogenólisis: Promueve la liberación de glucosa a partir del glucógeno almacenado en el
músculo.
3. Acciones en el Tejido Adiposo:
• Lipólisis: Estimula la descomposición de triglicéridos en ácidos grasos y glicerol, liberando
estos componentes para su uso como fuente de energía.
1. El glucagón estimula la lipólisis en el tejido adiposo al activar la lipasa sensible a
hormonas.
2. Indirectamente, el glucagón favorece la cetogénesis al aumentar la actividad de la
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
carnitina acetiltransferasa I (CAT I), que facilita la transferencia de ácidos grasos a las
mitocondrias para su oxidación.
3. La inhibición de la acetil-CoA carboxilasa por el glucagón reduce la concentración de
malonil-CoA, permitiendo así que la CAT I sea activada.
4. Acciones Extrahepáticas (en concentraciones farmacológicas):
• En condiciones fisiológicas, el hígado es el principal tejido diana del glucagón. Sin embargo,
en concentraciones farmacológicas, el glucagón también puede tener acciones en otros
tejidos, como el músculo cardíaco y esquelético, así como acciones lipolíticas y proteolíticas
en varios tejidos.
Secreción y Procesamiento del Glucagón:
• El glucagón es un péptido de 31 aminoácidos sintetizado por las células α de los islotes de
Langerhans en el páncreas.
• El gen del glucagón se encuentra en el cromosoma 2 y se traduce en preproglucagón.
• Durante la traducción del ARNm en el retículo endoplásmico rugoso, se elimina el péptido señal,
generando proglucagón.
• En las células α, el proglucagón es escindido en la molécula madura de glucagón y varios péptidos
biológicamente activos.
• Las células L intestinales procesan el proglucagón de manera diferente, produciendo GLP-1 y GLP-
2, que tienen acciones biológicas específicas. GLP-1 es una potente incretina que estimula la
secreción de insulina. El GLP-2 no es una incretina, y se desconocen sus acciones biológicas.
Regulación de la Secreción:
• Los aminoácidos son los principales secretagogos del glucagón, pero las concentraciones necesarias
para la secreción in vitro son mayores que las requeridas in vivo. Se sugiere que factores neurales o
humorales amplifican la respuesta in vivo.
• La glucosa y varios aminoácidos estimulan la secreción de insulina, pero solo los aminoácidos
estimulan la secreción de glucagón. La glucosa, por el contrario, inhibe la secreción de glucagón.
• Aunque el glucagón es un secretagogo de la insulina, su efecto paracrino sobre la secreción de
insulina es limitado debido a la ubicación de las células α en la cascada sanguínea; la mayoría de las
células α se localizan más abajo en la cascada sanguínea que las células β, es poco probable que el
glucagón ejerza un efecto paracrino importante sobre la secreción de insulina.
Interacción con el Receptor y Activación de Vías de Señalización:
• El glucagón ejerce su acción al unirse a su receptor, que activa la proteína G heterotrimérica Gαs.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
• La activación de Gαs estimula la adenilato-ciclasa, que convierte el ATP en AMPc.
• El AMPc resultante activa la proteína quinasa dependiente de AMPc (PKA), que fosforila diversas
enzimas y sustratos proteicos.
Regulación Negativa de Enzimas Clave por la Insulina:
• La insulina tiene efectos opuestos a los del glucagón en varias vías metabólicas hepáticas.
• La insulina induce la desfosforilación de ciertas enzimas clave, como la glucógeno-sintasa y la
fosforilasa, inhibiendo la glucogenólisis.
• La insulina limita la transcripción de enzimas gluconeogénicas y de la oxidación de ácidos grasos a
través de la vía PI3K/Akt.
Coordinación de Respuestas Metabólicas:
• El glucagón y la insulina trabajan en conjunto para mantener la homeostasis metabólica.
• En situaciones de ayuno, el glucagón estimula la producción hepática de glucosa y la
gluconeogénesis, proporcionando así combustible al cerebro y otros tejidos.
• La insulina, por otro lado, favorece el almacenamiento de glucosa y lípidos en el hígado y músculo
después de la ingestión de alimentos.
9.3.- Somatostatina.
• La somatostatina se produce en las células δ de los islotes pancreáticos, células D del tubo digestivo,
en el hipotálamo y en otras partes del sistema nervioso central.
• Existen formas de somatostatina de 14 y 28 aminoácidos. La forma de 14 aminoácidos es la porción
C-terminal de la forma de 28 aminoácidos. Ambas formas tienen actividad biológica, pero la forma
de 14 aminoácidos predomina en el hipotálamo, mientras que la forma de 28 aminoácidos predomina
en el tubo digestivo y en las células δ del páncreas.
Acciones Inhibitorias de la Somatostatina:
• La somatostatina ejerce efectos inhibitorios sobre la secreción de varias hormonas, incluyendo la
hormona del crecimiento, insulina, glucagón, gastrina, péptido intestinal vasoactivo (VIP) y hormona
estimulante de la tiroides.
• En el páncreas, la somatostatina inhibe la secreción basal de insulina.
Uso Terapéutico:
• Un análogo de la somatostatina de acción prolongada, llamado octreotida, se utiliza terapéuticamente
en tumores endocrinos difíciles de tratar, como aquellos que producen hormona del crecimiento
(acromegalia), insulina (insulinoma) o serotonina (carcinoide), entre otros.
9.4.- Diabetes Tipo I y Tipo II.
Origen:
• Diabetes Tipo 1 (DM1): Es una enfermedad autoinmune en la cual el sistema inmunológico ataca y
destruye las células beta del páncreas, que son responsables de producir insulina. Esto conduce a una
deficiencia total de insulina.
• Diabetes Tipo 2 (DM2): Generalmente se desarrolla más tarde en la vida y está asociada con la
resistencia a la insulina y la disminución de la secreción de insulina por parte de las células beta. La
autoinmunidad no es un componente clave en la diabetes tipo 2.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Insulina:
• DM1: Hay una deficiencia absoluta de insulina; los pacientes con diabetes tipo 1 dependen de la
administración externa de insulina para sobrevivir; los niveles de insulina en sangre son bajos o
indetectables.
• DM2: Puede haber resistencia a la insulina, pero en las etapas iniciales, los niveles de insulina en
sangre pueden ser normales o incluso elevados. Con el tiempo, la producción de insulina puede
disminuir. Con el tiempo, la producción de insulina puede disminuir, pero no al punto de la
deficiencia total observada en la diabetes tipo 1.
Edad de Aparición:
• DM1: A menudo se diagnostica en la infancia o adolescencia, pero puede ocurrir a cualquier edad.
• DM2: Se desarrolla generalmente en adultos, pero ahora también se observa cada vez más en
adolescentes y niños, especialmente debido a las tasas crecientes de obesidad.
Predisposición Genética:
• DM1: Tiene una fuerte predisposición genética. La presencia de ciertos marcadores genéticos puede
aumentar el riesgo de desarrollar diabetes tipo 1.
• DM2: También hay una predisposición genética, pero factores ambientales, como la obesidad y el
estilo de vida, desempeñan un papel más destacado en su desarrollo.
Tratamiento:
• DM1: Requiere insulina exógena como parte integral del tratamiento. Se utilizan inyecciones de
insulina o bombas de insulina.
• DM2: Inicialmente, puede manejarse con cambios en el estilo de vida, dieta y medicamentos orales.
En algunos casos, la insulina puede ser necesaria, especialmente a medida que progresa la
enfermedad.
Factores de Riesgo:
• DM1: Factores de riesgo menos claros, pero la genética y posiblemente algunos factores ambientales
desencadenantes pueden estar involucrados.
• DM2: Factores de riesgo bien establecidos incluyen la obesidad, la falta de actividad física, la edad
avanzada y la historia familiar de diabetes.
Síntomas Iniciales:
• DM1: Los síntomas suelen aparecer de manera abrupta y pueden incluir sed excesiva, micción
frecuente, pérdida de peso inexplicada, fatiga extrema y cambios en la visión. La cetoacidosis
metabólica es una complicación grave que puede ocurrir, llevando a síntomas como náuseas,
vómitos, respiración rápida y aliento con olor a frutas (debido a la presencia de cuerpos cetónicos).
• DM2: Los síntomas pueden desarrollarse de manera más gradual. Pueden incluir los mismos
síntomas que en la DM1, pero también es común que los pacientes no presenten síntomas notables en
las etapas iniciales.
Síndrome Metabólico:
• DM2: La diabetes tipo 2 está fuertemente asociada con el síndrome metabólico, que es un conjunto
de condiciones que incluyen resistencia a la insulina, obesidad abdominal, hipertensión arterial y
niveles elevados de triglicéridos y colesterol LDL, y bajos niveles de colesterol HDL. Estos factores
aumentan el riesgo de enfermedad cardiovascular.
• DM1: Aunque los pacientes con DM1 también pueden tener factores de riesgo cardiovascular, el
síndrome metabólico no es una característica distintiva de la DM1.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
Cetonas y Cetoacidosis:
• DM1: La cetoacidosis diabética es más común en la diabetes tipo 1. Se produce cuando los niveles
de insulina son muy bajos y el cuerpo comienza a descomponer las grasas para obtener energía, lo
que resulta en la acumulación de cuerpos cetónicos en la sangre.
• DM2: La cetoacidosis es rara en la diabetes tipo 2, ya que generalmente hay algo de insulina presente
para prevenir una descomposición significativa de las grasas.
Progresión de la Enfermedad:
• DM1: La progresión es generalmente rápida, y la insulina es necesaria desde el principio.
• DM2: La progresión puede ser lenta y gradual. Al principio, puede manejarse con cambios en el
estilo de vida y medicamentos orales, pero con el tiempo, la insulina puede ser necesaria.
FISIOLOGÍA MEDICA I | AMADEO CABELLO PÉREZ
También podría gustarte
- TALLER 3 - Fisiología Del Tiroides - 2020-1Documento10 páginasTALLER 3 - Fisiología Del Tiroides - 2020-1Laura C. Quintero AlvaradoAún no hay calificaciones
- Hormonas Tiroideas Reporte 5fv1Documento17 páginasHormonas Tiroideas Reporte 5fv1Said MossoAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Tiroides ExpoDocumento33 páginasTiroides ExpoNiebla AbelAún no hay calificaciones
- Farmacologia Hormona Tiroidea-1Documento70 páginasFarmacologia Hormona Tiroidea-1Edson Machicado VargasAún no hay calificaciones
- Tiroidesy Paratiroides, Practica de Endocrino. Onice LugoDocumento13 páginasTiroidesy Paratiroides, Practica de Endocrino. Onice LugoOnice LugoAún no hay calificaciones
- Seminario - Hormonas Tiroideas-1Documento14 páginasSeminario - Hormonas Tiroideas-1JOSUE PALMA RUIZAún no hay calificaciones
- Glándula tiroides: hormonas T3 y T4 regulan metabolismoDocumento4 páginasGlándula tiroides: hormonas T3 y T4 regulan metabolismoPaula SantanaAún no hay calificaciones
- 3.tiroides Naturales 2023Documento7 páginas3.tiroides Naturales 2023alejandro barnett matamorosAún no hay calificaciones
- Capítulo 36.Documento40 páginasCapítulo 36.Leonel Gonthier100% (1)
- Glándula Tiroides y ParatiroidesDocumento20 páginasGlándula Tiroides y ParatiroidesAlejandra LujánAún no hay calificaciones
- Fisiopatología del eje tiroidesDocumento13 páginasFisiopatología del eje tiroidesPotencia SalasAún no hay calificaciones
- Paciente TiroideoDocumento23 páginasPaciente TiroideoAndrea Elizabeth Ruíz HérculesAún no hay calificaciones
- Clase de TiroidesDocumento111 páginasClase de TiroidesLinda LadyAún no hay calificaciones
- ENDOCRINOFISIOLOGÍA Tiroides y ParatiroidesDocumento36 páginasENDOCRINOFISIOLOGÍA Tiroides y ParatiroidesLucero Itala Samame LauraAún no hay calificaciones
- Glandula Tiroides..ConfDocumento27 páginasGlandula Tiroides..ConfCristiane OliveiraAún no hay calificaciones
- TiroidesDocumento137 páginasTiroidesFernando Gonzalez S.Aún no hay calificaciones
- Ensayo de La TiroidesDocumento18 páginasEnsayo de La TiroidesnallelylunapalacioAún no hay calificaciones
- Resumen CEMVDocumento43 páginasResumen CEMVLaura Sanchez AguirreAún no hay calificaciones
- Anatomía Tema 10Documento4 páginasAnatomía Tema 10Cesar CubasAún no hay calificaciones
- MEDICINA HUMANA: Resumen de Fisiología de La Glándula TiroidesDocumento6 páginasMEDICINA HUMANA: Resumen de Fisiología de La Glándula Tiroidesreed lunasAún no hay calificaciones
- T3 y T4Documento6 páginasT3 y T4billyAún no hay calificaciones
- Hormona TiroideaDocumento23 páginasHormona TiroideaHOLGER IVAN RIVERA DELGADOAún no hay calificaciones
- II.1 Función Tiroidea - Hormonas TiroideasDocumento200 páginasII.1 Función Tiroidea - Hormonas TiroideasMauricio VidalAún no hay calificaciones
- FisioexamenDocumento15 páginasFisioexamenMarié AguilarAún no hay calificaciones
- Anatomía y Fisiología Del Eje Hipotálamo - Hipófisis - TiroidesDocumento3 páginasAnatomía y Fisiología Del Eje Hipotálamo - Hipófisis - TiroidesMaría Fernanda Hurtado B.Aún no hay calificaciones
- Patología de La Glándula TiroidesDocumento36 páginasPatología de La Glándula TiroidesCarolina OrdoñezAún no hay calificaciones
- HORMONAS TIROIDEAS - Modo de CompatibilidadDocumento208 páginasHORMONAS TIROIDEAS - Modo de CompatibilidadAnalitab Unidad de DiagnosticoAún no hay calificaciones
- 3.2 Estudio Bioquímico de La Función TiroideaDocumento61 páginas3.2 Estudio Bioquímico de La Función TiroideaInés VelardeAún no hay calificaciones
- Fisiología Del Hipotiroidismo en AnimalesDocumento14 páginasFisiología Del Hipotiroidismo en AnimalesAlesu RodríguezAún no hay calificaciones
- Asi Somos El Cuerpo Humano F22 - 2023Documento8 páginasAsi Somos El Cuerpo Humano F22 - 2023Edgar GuaitaAún no hay calificaciones
- Tiroides y Paratiroides-2Documento11 páginasTiroides y Paratiroides-2salejandra03Aún no hay calificaciones
- Apuntes EndocrinoDocumento197 páginasApuntes EndocrinoMishel VillaAún no hay calificaciones
- Farmacología de Las Hormonas TiroideasDocumento23 páginasFarmacología de Las Hormonas TiroideasNoelia GomezAún no hay calificaciones
- Siste EndocrinoDocumento29 páginasSiste EndocrinoSergio AguileraAún no hay calificaciones
- Medicina - Fisiologia. Hormonas TiroideasDocumento23 páginasMedicina - Fisiologia. Hormonas TiroideasVic_jvpAún no hay calificaciones
- Tema 1 - Tiroides, Hipotiroidismo y Coma Mixedematoso.Documento14 páginasTema 1 - Tiroides, Hipotiroidismo y Coma Mixedematoso.Medicina 5 Seccion 11Aún no hay calificaciones
- Hipotiroidismo SemioDocumento29 páginasHipotiroidismo SemioPierePreciadoCumpaAún no hay calificaciones
- Trabajo Colaborativo Hormonas Tiroideas Fisiologia I - III Semestre - 2023-1Documento7 páginasTrabajo Colaborativo Hormonas Tiroideas Fisiologia I - III Semestre - 2023-1María VarónAún no hay calificaciones
- Farmacología de Las Hormonas Tiroideas LPLR 3FDocumento5 páginasFarmacología de Las Hormonas Tiroideas LPLR 3FKari R AgredaAún no hay calificaciones
- Sintesis de Hormona TiroideaDocumento9 páginasSintesis de Hormona Tiroideagustavo manzanedo carbajalAún no hay calificaciones
- Anatomía y fisiología de la glándula tiroides (Anatomy and physiology of the thyroid glandDocumento4 páginasAnatomía y fisiología de la glándula tiroides (Anatomy and physiology of the thyroid glandLeslie Thalia Rojas JustinianoAún no hay calificaciones
- Formación de La Hormona TiroideaDocumento4 páginasFormación de La Hormona TiroideaPaolaAún no hay calificaciones
- Glándula TiroidesDocumento26 páginasGlándula TiroideskennethAún no hay calificaciones
- Hormonas Metabolicas TiroideasDocumento6 páginasHormonas Metabolicas TiroideasjyanesAún no hay calificaciones
- TiroidesDocumento51 páginasTiroidesluna.zamudio.angelicasamantha.21Aún no hay calificaciones
- TiroidesDocumento8 páginasTiroidesIsi AlvarezAún no hay calificaciones
- Práctica 7 FisiologíaDocumento33 páginasPráctica 7 FisiologíaGerson GarciaAún no hay calificaciones
- Fisiología Endocrina 2Documento69 páginasFisiología Endocrina 2Analy AstupiñanAún no hay calificaciones
- Fármacos tiroideos y antitiroideosDocumento4 páginasFármacos tiroideos y antitiroideosFRANCISCO NARANJOAún no hay calificaciones
- Glándula TiroideaDocumento11 páginasGlándula TiroideaMaguita ArmijosAún no hay calificaciones
- Función hormonal de la glándula tiroidesDocumento24 páginasFunción hormonal de la glándula tiroidesJN KAún no hay calificaciones
- Cuestionario EndocrinoDocumento6 páginasCuestionario Endocrinomaria judith ugarte guevaraAún no hay calificaciones
- Farmacologia 28Documento11 páginasFarmacologia 28R Martín Apresa MAún no hay calificaciones
- hormonas-metabolicas-tiroidesDocumento16 páginashormonas-metabolicas-tiroidesnvallejos681Aún no hay calificaciones
- Fisiología de La TiroidesDocumento38 páginasFisiología de La TiroidesMayerly MartinezAún no hay calificaciones
- BiologiaDocumento7 páginasBiologiaMarck RosseAún no hay calificaciones
- Grandula TiroidesDocumento2 páginasGrandula TiroidesPatricia UrbinaAún no hay calificaciones
- Acidos nucleicos y nucleótidosDocumento46 páginasAcidos nucleicos y nucleótidosG.CAún no hay calificaciones
- Zinc óxido propiedades y usosDocumento2 páginasZinc óxido propiedades y usosLeonardo LozanoAún no hay calificaciones
- TALLER Filosofos de La NaturalezaDocumento4 páginasTALLER Filosofos de La NaturalezaANGELICA HERNANDEZ ESCOBARAún no hay calificaciones
- Diseño y Construcción de Robot EscaladorDocumento145 páginasDiseño y Construcción de Robot EscaladorUnitronik Robotica0% (1)
- Notación AlgebraicaDocumento2 páginasNotación Algebraicaangelito131Aún no hay calificaciones
- Vinno E10Documento4 páginasVinno E10darmanex.sasAún no hay calificaciones
- Cuidados EPOCDocumento5 páginasCuidados EPOCConstanza Abalos RodríguezAún no hay calificaciones
- Cultivo y Proceso Agroindustrial de Zacate LimónDocumento34 páginasCultivo y Proceso Agroindustrial de Zacate LimónMelvin GarciaAún no hay calificaciones
- Tesis FINAL 23eneroDocumento161 páginasTesis FINAL 23eneroRuben MataAún no hay calificaciones
- Informe 7 Comunicaciones AnalogicasDocumento4 páginasInforme 7 Comunicaciones AnalogicasSANTIAGO PARRA TOROAún no hay calificaciones
- Cultivo Faringeo1Documento16 páginasCultivo Faringeo1Anneth Michel Salinas GastelumAún no hay calificaciones
- Placa Dentobacteriana Articulo PDFDocumento4 páginasPlaca Dentobacteriana Articulo PDFTirsa GonzalezAún no hay calificaciones
- 2.8L diesel motor manual KJDocumento107 páginas2.8L diesel motor manual KJJorge Andres Fritis EstayAún no hay calificaciones
- Taller PresupuestosDocumento9 páginasTaller PresupuestosFresa giraldoAún no hay calificaciones
- DEMOLICIÓN DE BEBEDEROSDocumento1 páginaDEMOLICIÓN DE BEBEDEROSJorge Bañuelos100% (1)
- Cuenca Hidrografica Del PeruDocumento20 páginasCuenca Hidrografica Del PeruAlfredo Carpio TorresAún no hay calificaciones
- DPFDocumento4 páginasDPFDIONYBLINKAún no hay calificaciones
- Bendición y Consagración Del TemploDocumento12 páginasBendición y Consagración Del TemploDiegoAún no hay calificaciones
- Dinamica de Los Grupos Segun Kurt LewinDocumento3 páginasDinamica de Los Grupos Segun Kurt LewinCarolina MejiaAún no hay calificaciones
- Analisis de Fisica ModernaDocumento2 páginasAnalisis de Fisica ModernajuanAún no hay calificaciones
- Informe Museo Machupicchu 1991Documento51 páginasInforme Museo Machupicchu 1991don gato100% (1)
- Examen Final Quimica FotograficaDocumento3 páginasExamen Final Quimica FotograficaHipolito CruzAún no hay calificaciones
- Recuperación Entorno de Los TollosDocumento6 páginasRecuperación Entorno de Los TollosHoracio Gonzalez AndradesAún no hay calificaciones
- Las MitocondriasDocumento2 páginasLas Mitocondrias0ZKRCITHOAún no hay calificaciones
- Fisiología Animal Básica.Documento347 páginasFisiología Animal Básica.fernie3007Aún no hay calificaciones
- LAVADORAS Falla y SolucionesDocumento9 páginasLAVADORAS Falla y SolucionesMechín Rodriguez EjvrAún no hay calificaciones
- Foda de Residuos Solidos: Integrantes: 1. Vilca Montes, David 2. Chavez Curasma, Lizbeth 3. Escobar Ccanto, SandyDocumento4 páginasFoda de Residuos Solidos: Integrantes: 1. Vilca Montes, David 2. Chavez Curasma, Lizbeth 3. Escobar Ccanto, Sandysandy escobar ccantoAún no hay calificaciones
- El Procedimiento de Servidumbre MineraDocumento4 páginasEl Procedimiento de Servidumbre MineraWskacer Cerqos FaxseAún no hay calificaciones
- Serie 2Documento5 páginasSerie 2942107995Aún no hay calificaciones
- Bisoprolol: bloqueante beta1 altamente selectivoDocumento3 páginasBisoprolol: bloqueante beta1 altamente selectivoJOHA1408Aún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Fisiología renal y metabolismo hidrosalino: Segunda ediciónDe EverandFisiología renal y metabolismo hidrosalino: Segunda ediciónAún no hay calificaciones
- Mediciones y métodos de uso común en el laboratorio de QuímicaDe EverandMediciones y métodos de uso común en el laboratorio de QuímicaCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Fundamentos de química aplicados a las ciencias de la saludDe EverandFundamentos de química aplicados a las ciencias de la saludAún no hay calificaciones
- Curso Práctico de Química GeneralDe EverandCurso Práctico de Química GeneralCalificación: 4 de 5 estrellas4/5 (4)
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Bioquímica: metabolismo energético, conceptos y aplicaciónDe EverandBioquímica: metabolismo energético, conceptos y aplicaciónCalificación: 4 de 5 estrellas4/5 (7)
- Sistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaDe EverandSistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaAún no hay calificaciones
- La química al alcance de todosDe EverandLa química al alcance de todosCalificación: 1 de 5 estrellas1/5 (1)
- Fisiología comparada del medio internoDe EverandFisiología comparada del medio internoCalificación: 3 de 5 estrellas3/5 (1)
- Las cadenas fisiológicas (Tomo II): La cintura pélvica y el miembro inferiorDe EverandLas cadenas fisiológicas (Tomo II): La cintura pélvica y el miembro inferiorCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Polímeros: Generalidades y tendencias de investigación en ColombiaDe EverandPolímeros: Generalidades y tendencias de investigación en ColombiaCalificación: 5 de 5 estrellas5/5 (14)
- Inteligencia física: Aprende a ver el mundo con la mente de un físicoDe EverandInteligencia física: Aprende a ver el mundo con la mente de un físicoCalificación: 5 de 5 estrellas5/5 (41)
- Estructura atómica y enlace químicoDe EverandEstructura atómica y enlace químicoCalificación: 3 de 5 estrellas3/5 (2)
- Problemas resueltos de Química Analítica CuantitativaDe EverandProblemas resueltos de Química Analítica CuantitativaCalificación: 5 de 5 estrellas5/5 (3)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Nomenclatura de las sustancias químicasDe EverandNomenclatura de las sustancias químicasCalificación: 5 de 5 estrellas5/5 (2)
- Sobre el combate: La psicología y fisiología del conflicto letal en la guerra y en la pazDe EverandSobre el combate: La psicología y fisiología del conflicto letal en la guerra y en la pazCalificación: 4 de 5 estrellas4/5 (9)
- Dieta para el Hipotiroidismo: Recetas para curar el hipotiroidismo, el hipertiroidismo y bajar de peso rápido: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandDieta para el Hipotiroidismo: Recetas para curar el hipotiroidismo, el hipertiroidismo y bajar de peso rápido: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (3)
- Aspectos claves Tiroides: Primer EdiciónDe EverandAspectos claves Tiroides: Primer EdiciónCalificación: 5 de 5 estrellas5/5 (4)