También podría gustarte
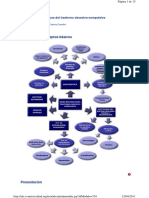
- Neonatos y lactantes menores: Guía orofacial y deglutoria. Evaluación, diagnóstico e intervención terapéuticaDe EverandNeonatos y lactantes menores: Guía orofacial y deglutoria. Evaluación, diagnóstico e intervención terapéuticaCalificación: 4 de 5 estrellas4/5 (6)
- RODILLADocumento62 páginasRODILLAeliseoo8ortega100% (1)
- Desarrollo cognitivo en niños prematuros y de bajo peso al nacerDe EverandDesarrollo cognitivo en niños prematuros y de bajo peso al nacerAún no hay calificaciones
- Manual CTO de Medicina y Cirugía - UG (Urgencias y Medicina Intensiva) - 12° Edición (2021)Documento28 páginasManual CTO de Medicina y Cirugía - UG (Urgencias y Medicina Intensiva) - 12° Edición (2021)JoseLuis0% (1)
- Depresion Post Parto Aspectos Clinicos PDFDocumento11 páginasDepresion Post Parto Aspectos Clinicos PDFCarla Fernandez100% (1)
- TDAHDocumento21 páginasTDAHTania Muchica PizarroAún no hay calificaciones
- Examen de Aprobación Del Curso de Transporte de Materiales PeligrososDocumento6 páginasExamen de Aprobación Del Curso de Transporte de Materiales PeligrososMarco Fernando Bravo50% (2)
- Teoria de La Mente I v.2Documento13 páginasTeoria de La Mente I v.2Yuneydis GuzmanAún no hay calificaciones
- Test 592 y DiamantesDocumento2 páginasTest 592 y DiamantesAnaaaerobios100% (1)
- Marco Teorico Tdah, Tda y TNDDocumento17 páginasMarco Teorico Tdah, Tda y TNDYamiAún no hay calificaciones
- Marco Teorico - TDHDocumento8 páginasMarco Teorico - TDHXiomara RodriguezAún no hay calificaciones
- Causas TdahDocumento2 páginasCausas TdahAngelica Maria Santana CoronadoAún no hay calificaciones
- Cognicion y Teorias Del DesarrolloDocumento33 páginasCognicion y Teorias Del DesarrolloEmily Vega ArdilaAún no hay calificaciones
- Autismo NewDocumento3 páginasAutismo NewArnel SmitAún no hay calificaciones
- CAPÍTULO 23 - Trastorno FetalDocumento5 páginasCAPÍTULO 23 - Trastorno FetalandreaAún no hay calificaciones
- Trastornos Del Desarrollo en Niños y AdolescentesDocumento18 páginasTrastornos Del Desarrollo en Niños y AdolescentesYISUS MAAún no hay calificaciones
- Marc Teor PDFDocumento5 páginasMarc Teor PDFroberto gomezAún no hay calificaciones
- Tdah - Matriz de ConsistenciaDocumento4 páginasTdah - Matriz de ConsistenciaFanyta AtaramaAún no hay calificaciones
- 389209-Texto Del Artículo-563378-1-10-20210624Documento2 páginas389209-Texto Del Artículo-563378-1-10-20210624Eleanor AlmAún no hay calificaciones
- 5 Sesión Problemas de AprendizajeDocumento3 páginas5 Sesión Problemas de AprendizajeLiliana MarceloAún no hay calificaciones
- HIPERACTIVIDADDocumento54 páginasHIPERACTIVIDADSilvia Galindo MartinezAún no hay calificaciones
- Alteraciones Neuropsiquiátricas en Menores de 18 Años Expuestos A Opioides en El Periodo Prenatal: Revisión NarrativaDocumento9 páginasAlteraciones Neuropsiquiátricas en Menores de 18 Años Expuestos A Opioides en El Periodo Prenatal: Revisión NarrativaCarolina GuzmánAún no hay calificaciones
- Autismo Equio 123Documento32 páginasAutismo Equio 123Nallely P. IbarraAún no hay calificaciones
- Trastorno de Deficit de Atencion e Hiperactividad (TDAH)Documento17 páginasTrastorno de Deficit de Atencion e Hiperactividad (TDAH)GoNzalita D DiOs GonzalezAún no hay calificaciones
- Resumen TdahDocumento19 páginasResumen TdahSalvador HUEBRA ARANAAún no hay calificaciones
- Factores de Riesgos.Documento7 páginasFactores de Riesgos.Maria Jose Barraza CAún no hay calificaciones
- Marco TeoricoDocumento2 páginasMarco TeoricoFelipe MuñozAún no hay calificaciones
- Etiología de Los Trastornos de La AtenciónDocumento4 páginasEtiología de Los Trastornos de La Atenciónjosephine martinez marinAún no hay calificaciones
- Los Trastornos de La Conducta AlimentariaDocumento5 páginasLos Trastornos de La Conducta Alimentarialauradiana.vsAún no hay calificaciones
- Avance TOCDocumento8 páginasAvance TOCEli RomanAún no hay calificaciones
- Wuolah Free TDAHDocumento11 páginasWuolah Free TDAHAndreaAún no hay calificaciones
- TDAH y TtoDocumento29 páginasTDAH y TtoGuillermo Chacon AcevedoAún no hay calificaciones
- Etiologia Del TdahDocumento3 páginasEtiologia Del TdahJuan IsazaAún no hay calificaciones
- Cuerpo TDAHDocumento10 páginasCuerpo TDAHLuffy MugiwaraAún no hay calificaciones
- AutismoDocumento4 páginasAutismoLaura Marcela Baldrich CorreaAún no hay calificaciones
- TDAHDocumento14 páginasTDAHFanahí González LópAún no hay calificaciones
- Ps123 03Documento6 páginasPs123 03Doego BarreraAún no hay calificaciones
- La Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo EmbrionarioDocumento10 páginasLa Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo Embrionariocamila gonzalezAún no hay calificaciones
- Articulo Efectos Del Estres Materno Durante El Embarazo y Su Repercusion Sobre El Neurodesarrollo Del FetoDocumento6 páginasArticulo Efectos Del Estres Materno Durante El Embarazo y Su Repercusion Sobre El Neurodesarrollo Del FetoLORETH VALERIA GOMEZ GOMEZAún no hay calificaciones
- Depresión Materna y Vínculo Madre-BebéDocumento22 páginasDepresión Materna y Vínculo Madre-BebéTruelost BlqvqAún no hay calificaciones
- Trabajo Final VanessaDocumento37 páginasTrabajo Final VanessaJulio HuancaAún no hay calificaciones
- Programación en Etapas Tempranas de La VidaDocumento17 páginasProgramación en Etapas Tempranas de La VidaDianajas garciaAún no hay calificaciones
- Factores Que Se Relacionan Con La Depresión en Esta EtapaDocumento4 páginasFactores Que Se Relacionan Con La Depresión en Esta EtapaCuori JsaAún no hay calificaciones
- Ensayo TdaDocumento9 páginasEnsayo TdaNalleli Sanchez FloresAún no hay calificaciones
- Maldonado-Duran-El Por Que y Como Salud Mental BebeDocumento10 páginasMaldonado-Duran-El Por Que y Como Salud Mental BebePaulina Andrea Alvarez UrrutiaAún no hay calificaciones
- TM en GinecoDocumento9 páginasTM en GinecoArias Gutierrez KatherineAún no hay calificaciones
- Trabajo FinalDocumento8 páginasTrabajo FinalKelly Del Rosario Salazar YariceAún no hay calificaciones
- Tdah Correcciones y Entrega 6o%Documento17 páginasTdah Correcciones y Entrega 6o%アルダナ クラウディアAún no hay calificaciones
- Es El Autismo Una Enfermedad Del CerebeloDocumento45 páginasEs El Autismo Una Enfermedad Del CerebeloLidaConstanzaMendezAún no hay calificaciones
- CAPÍTULO 1 - BoorrDocumento6 páginasCAPÍTULO 1 - Boorrfern392Aún no hay calificaciones
- Trastornos MentalesDocumento15 páginasTrastornos MentalesCARO CHAMORROAún no hay calificaciones
- 4-Efectos Del Estrés Materno Intenso y Prolongado Durante El Embarazo y SuDocumento7 páginas4-Efectos Del Estrés Materno Intenso y Prolongado Durante El Embarazo y Suvarios0215Aún no hay calificaciones
- Los Trastornos Generaido en Paginas - AlcasDocumento14 páginasLos Trastornos Generaido en Paginas - AlcasJosé NatsuharaAún no hay calificaciones
- Carga Alostática en Trastorno BipolarDocumento14 páginasCarga Alostática en Trastorno BipolarAMCArroyo100% (1)
- Bases Biologicas Del Trastorno Obsesivo-CompulsivoDocumento15 páginasBases Biologicas Del Trastorno Obsesivo-CompulsivoBunburys BarAún no hay calificaciones
- Javier & Vallejo TdahDocumento8 páginasJavier & Vallejo TdahMonica LeonorAún no hay calificaciones
- Javier & Vallejo TdahDocumento8 páginasJavier & Vallejo TdahMonica LeonorAún no hay calificaciones
- Trastorno Por Déficit de Atención Con Hiperactividad PDFDocumento14 páginasTrastorno Por Déficit de Atención Con Hiperactividad PDFKiko ChevarriaAún no hay calificaciones
- Content ServerDocumento10 páginasContent ServerJuan David Bermudez MoralesAún no hay calificaciones
- Tdah TesisDocumento19 páginasTdah TesisAngelica Esquivel TreviñoAún no hay calificaciones
- 02 Modulo 2 TDAHDocumento17 páginas02 Modulo 2 TDAHAnna Canet TarrésAún no hay calificaciones
- Deficit Atencion Hiperactividad PedagogicoDocumento22 páginasDeficit Atencion Hiperactividad PedagogicomareanidAún no hay calificaciones
- Ganglios BasalesDocumento10 páginasGanglios BasalesSofia Fernandez LuqueAún no hay calificaciones
- Tdah Investigacion 3Documento13 páginasTdah Investigacion 3Carolina Castillo100% (1)
- Neuropsicologia - y - Esquizofrenia - Cuesta (2000)Documento13 páginasNeuropsicologia - y - Esquizofrenia - Cuesta (2000)AnaaaerobiosAún no hay calificaciones
- Partida Nacimieto AbueloDocumento1 páginaPartida Nacimieto AbueloLidia LazzaroAún no hay calificaciones
- Cómo Preparar Un Latte de Vainilla de StarbucksDocumento3 páginasCómo Preparar Un Latte de Vainilla de StarbucksAnaaaerobiosAún no hay calificaciones
- Dieta de ScardaleDocumento4 páginasDieta de ScardaleAnaaaerobiosAún no hay calificaciones
- Consonant Trigrams Test (CTT) (Brown & Peterson, 1958) : Estímulos Comenzar Número Delay (Seg.) RespuestasDocumento1 páginaConsonant Trigrams Test (CTT) (Brown & Peterson, 1958) : Estímulos Comenzar Número Delay (Seg.) RespuestasAnaaaerobios100% (1)
- Denver Test SimplificadoDocumento14 páginasDenver Test SimplificadoAnaaaerobiosAún no hay calificaciones
- Nuestra Opinión Acerca de Nosotros Mismos y Del MundoDocumento10 páginasNuestra Opinión Acerca de Nosotros Mismos y Del MundoMaria Ignacia GonzalezAún no hay calificaciones
- Programa Diploma Salud OcupacionalDocumento10 páginasPrograma Diploma Salud OcupacionalLuis-alfredo Perez Bolde HernandezAún no hay calificaciones
- NORMASDocumento45 páginasNORMASMilena ObandoAún no hay calificaciones
- Refrigeracion y CongelacionDocumento9 páginasRefrigeracion y CongelacionNoemi Canales LeonAún no hay calificaciones
- Pineda Delgado, Ensayo, Grupo 101Documento2 páginasPineda Delgado, Ensayo, Grupo 101MIA FERNANDA PINEDA DELGADOAún no hay calificaciones
- 0 - Criterio Diagnóstico de Dislexia y DisgrafiaDocumento1 página0 - Criterio Diagnóstico de Dislexia y DisgrafiaSergio polo yepesAún no hay calificaciones
- Com Ecom RecibodigitalDocumento1 páginaCom Ecom RecibodigitalLuis Miguel BuksaAún no hay calificaciones
- Plan Charla ClínicaDocumento2 páginasPlan Charla ClínicaLupitha PMAún no hay calificaciones
- Como Hacer Examen CoprologicoDocumento11 páginasComo Hacer Examen CoprologicoDaniela CepedaAún no hay calificaciones
- Relación CéntricaDocumento3 páginasRelación CéntricaAndrea BaezAún no hay calificaciones
- IL - Limpieza y Desinfección de JaulasDocumento2 páginasIL - Limpieza y Desinfección de JaulasSofia CbaAún no hay calificaciones
- Orientaciones Metodológicas para La EnseñanzaDocumento4 páginasOrientaciones Metodológicas para La EnseñanzaBartolomé Natalia SabrinaAún no hay calificaciones
- Alexander Cancino MoralesDocumento4 páginasAlexander Cancino MoralesAlexander CancinoAún no hay calificaciones
- Uc2 - 02H - Sesion 14 - 2021Documento12 páginasUc2 - 02H - Sesion 14 - 2021Edgar Quispe AlfaroAún no hay calificaciones
- Artículos de Interés Sobre FitoterapiaDocumento85 páginasArtículos de Interés Sobre FitoterapiasanpedrolagunillasAún no hay calificaciones
- Enfermedades de OceaniaDocumento1 páginaEnfermedades de OceaniaDaniela MoralesAún no hay calificaciones
- Informe Empresa Quality ProductsDocumento30 páginasInforme Empresa Quality ProductsJHONATAN DANIEL CRUZADO LEONAún no hay calificaciones
- Propuesta Promocion y Prevencion en Salud MentalDocumento3 páginasPropuesta Promocion y Prevencion en Salud MentalCAROLINA QUIJANOAún no hay calificaciones
- Extraccion Denticion Primaria (INFANTIL)Documento4 páginasExtraccion Denticion Primaria (INFANTIL)Edna Lizette Vazquez AngelesAún no hay calificaciones
- Clase 2 - Apuntes Sobre El Proceso CreativoDocumento7 páginasClase 2 - Apuntes Sobre El Proceso CreativomarcomorenomunozAún no hay calificaciones
- Tema 1Documento48 páginasTema 1Elias Tarifa CondoriAún no hay calificaciones
- Proyecto 5S Grupo 1 (Ii)Documento52 páginasProyecto 5S Grupo 1 (Ii)Tefa AponteAún no hay calificaciones
- HS BacoxinDocumento4 páginasHS BacoxinPests Cleaning Cali - MedellinAún no hay calificaciones
- Indicadores de Salud Integral 18.09.20Documento8 páginasIndicadores de Salud Integral 18.09.20Jose Carlos HolgadoAún no hay calificaciones
- Final Guías Enfr. BásicaDocumento373 páginasFinal Guías Enfr. BásicaNaomy NavarreteAún no hay calificaciones
- Mapas Conceptuales: Catedrática: Ing. Maribel Montes Integrantes - Grupo B - Manejo de Desechos ComercialesDocumento45 páginasMapas Conceptuales: Catedrática: Ing. Maribel Montes Integrantes - Grupo B - Manejo de Desechos ComercialesHeymiAún no hay calificaciones
- Hombr DolorosoDocumento23 páginasHombr DolorosoStephanie SánchezAún no hay calificaciones