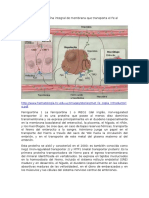
También podría gustarte
- Tra Ducci OnDocumento80 páginasTra Ducci OnDiego QuinterosAún no hay calificaciones
- Bioinorgánica Del HierroDocumento8 páginasBioinorgánica Del HierroJuan Wilfredo Hernandez Diaz75% (4)
- Repaso 1 Biología IB Año 1Documento5 páginasRepaso 1 Biología IB Año 1Juan Manuel Londoño50% (2)
- TEMA 1. Generalidades de Nutricion de BerriesDocumento88 páginasTEMA 1. Generalidades de Nutricion de BerriesJOSE MORA100% (1)
- Metabolismo Del HierroDocumento5 páginasMetabolismo Del Hierrogiovanna ramosAún no hay calificaciones
- Introducción Hierro Hem y No HemDocumento17 páginasIntroducción Hierro Hem y No HemYubissa MGAún no hay calificaciones
- Almacenamiento Del HierroDocumento2 páginasAlmacenamiento Del HierroJoshue MurrietaAún no hay calificaciones
- Ferropenia y Otras Anemia Hiporpoliferativas Versión BuenaDocumento11 páginasFerropenia y Otras Anemia Hiporpoliferativas Versión BuenaAmbar ToledoAún no hay calificaciones
- Anemia y Su ClasificacionDocumento7 páginasAnemia y Su ClasificacionAngela CVAún no hay calificaciones
- Cuestionario y VocabularioDocumento2 páginasCuestionario y VocabularioVilma Zeballos HuamaniAún no hay calificaciones
- Transferrin ADocumento8 páginasTransferrin Asuce_DAún no hay calificaciones
- Metabolismo Del HierroDocumento27 páginasMetabolismo Del HierroKarla López CAún no hay calificaciones
- Seminario Del HierroDocumento8 páginasSeminario Del HierroJuan Carlos Tenecora QuitoAún no hay calificaciones
- Cuestinario de Metabolismo de HierroDocumento4 páginasCuestinario de Metabolismo de HierroVane AguilarAún no hay calificaciones
- HEMOPROTEINASDocumento2 páginasHEMOPROTEINASVale DegraveAún no hay calificaciones
- Metabolismo de Fe, Anemia FerropénicaDocumento7 páginasMetabolismo de Fe, Anemia FerropénicaCatalina CirausAún no hay calificaciones
- Metabolismo Del HierroDocumento13 páginasMetabolismo Del HierroBelen DuceAún no hay calificaciones
- ¿Qué Es El Efecto Haldane?Documento4 páginas¿Qué Es El Efecto Haldane?Antony Barboza GalvezAún no hay calificaciones
- Metabolismo Del HierroDocumento47 páginasMetabolismo Del HierroNelson TellezAún no hay calificaciones
- Hierro y Cobre... para EstudiarDocumento12 páginasHierro y Cobre... para EstudiarLuis Felipe Giraldo LondoñoAún no hay calificaciones
- HIERRODocumento3 páginasHIERROWilliam EspinozaAún no hay calificaciones
- Metabolismo Del HierroDocumento27 páginasMetabolismo Del HierroLuis Antonio García RomeroAún no hay calificaciones
- Química Bioinorgánica Del HierroDocumento30 páginasQuímica Bioinorgánica Del HierroDiana De Fatima RomeroAún no hay calificaciones
- Preguntas de Metabolismo de HierroDocumento3 páginasPreguntas de Metabolismo de HierroAle Roque100% (1)
- Cofactores Enzimas (Metales)Documento12 páginasCofactores Enzimas (Metales)JOSE EAún no hay calificaciones
- Cofactores Enzimáticos Inorgánicos y OrganicosDocumento22 páginasCofactores Enzimáticos Inorgánicos y OrganicosAdolfo La Roche100% (1)
- HEPCIDINADocumento19 páginasHEPCIDINAFabi M. Quique PerezAún no hay calificaciones
- Metabolismo Del HierroDocumento18 páginasMetabolismo Del HierroHeriberto Richard IllanesAún no hay calificaciones
- Hierro PT.1Documento2 páginasHierro PT.1Yasuri Celia Trujillo GarcíaAún no hay calificaciones
- HierroDocumento5 páginasHierroFernando Cordova PardoAún no hay calificaciones
- FerroportinaDocumento4 páginasFerroportinaAriel Lòpez VelazquezAún no hay calificaciones
- Metabolismo Del HierroDocumento11 páginasMetabolismo Del HierroMario PerezAún no hay calificaciones
- Sem 6 Presencial HEMOGLOBINADocumento22 páginasSem 6 Presencial HEMOGLOBINACarlosAlbertoDangeloToledoAún no hay calificaciones
- Ciclo de HierroDocumento3 páginasCiclo de HierroXio Dklm0% (2)
- Fisio Pathos Del HierroDocumento52 páginasFisio Pathos Del HierrodZAún no hay calificaciones
- 3.el Hierro Es Un Elemento Esencial para La VidaDocumento7 páginas3.el Hierro Es Un Elemento Esencial para La VidaJasna ZovakAún no hay calificaciones
- Bioquímica Bloque II DefinitivoDocumento46 páginasBioquímica Bloque II Definitivoclaramed05Aún no hay calificaciones
- Pregunta 1Documento3 páginasPregunta 1Seydy Briggetty Tenazoa LopezAún no hay calificaciones
- Clase 7 Metabolismo Del Grupo HemoDocumento17 páginasClase 7 Metabolismo Del Grupo HemoAguss FernandezAún no hay calificaciones
- Metabolismo Del Hierro - Fisiologà - A IDocumento13 páginasMetabolismo Del Hierro - Fisiologà - A IKeka QuijanoAún no hay calificaciones
- Met HierroDocumento22 páginasMet HierroJuan Carlos AvendañoAún no hay calificaciones
- Anemia FerropenicaDocumento47 páginasAnemia FerropenicaCassandra TorresAún no hay calificaciones
- Metabolismo Del HierroDocumento13 páginasMetabolismo Del HierroRubby CsolAún no hay calificaciones
- V. Anemia Ferropenica y Sobrecarga de HierroDocumento49 páginasV. Anemia Ferropenica y Sobrecarga de HierroCitlali SanchezAún no hay calificaciones
- Bacterias Quimiolitótrofas Del Hierro y AzufreDocumento4 páginasBacterias Quimiolitótrofas Del Hierro y AzufreManu Jiménez García100% (3)
- Metabolismo Del HierroDocumento2 páginasMetabolismo Del HierroNickAún no hay calificaciones
- Metabolismo Del HierroDocumento3 páginasMetabolismo Del HierroDeyvis Escalante100% (1)
- Absorción y Metabolismo Del Hierro - MGVDocumento10 páginasAbsorción y Metabolismo Del Hierro - MGVMica Salas VictoriaAún no hay calificaciones
- Hemoglobina - HierroDocumento10 páginasHemoglobina - HierroEunice GrandaAún no hay calificaciones
- Monografia Anemia FerropenicaDocumento42 páginasMonografia Anemia FerropenicaDiego PacherresAún no hay calificaciones
- Metabolismo de HierroDocumento4 páginasMetabolismo de HierroJUAN RAMON PEREZ CONCEPCIONAún no hay calificaciones
- Estudio Del HierroDocumento24 páginasEstudio Del HierroPablo Ignacio Hernández RivasAún no hay calificaciones
- Hierro - 2 234Documento7 páginasHierro - 2 234karenAún no hay calificaciones
- ANEMIA FisiopatologiaDocumento51 páginasANEMIA Fisiopatologiaspartaco82Aún no hay calificaciones
- Metabolismo de HierroDocumento6 páginasMetabolismo de HierrofiorellaAún no hay calificaciones
- Metabolismo Del HierroDocumento2 páginasMetabolismo Del HierroGastón RodriguezAún no hay calificaciones
- Hierro MetabolismoDocumento18 páginasHierro MetabolismojuanetitoAún no hay calificaciones
- La TransferrinaDocumento3 páginasLa TransferrinaozimaxozaAún no hay calificaciones
- 03.003 Anemias CarencialesDocumento10 páginas03.003 Anemias CarencialesSabrina BronfenAún no hay calificaciones
- Metabolismo OxidativoDocumento21 páginasMetabolismo OxidativoPamelaSantaCruzRuizAún no hay calificaciones
- Metabolismo Del Hierro Folico y b12Documento19 páginasMetabolismo Del Hierro Folico y b12Remmy ValdezAún no hay calificaciones
- Discusión #3 de EnzimasDocumento7 páginasDiscusión #3 de EnzimasVanessa PortalAún no hay calificaciones
- Conversión de Lípidos en CarbohidratosDocumento7 páginasConversión de Lípidos en CarbohidratosCarlos Bonifaz67% (3)
- La Creatina Es Un PéptidoDocumento7 páginasLa Creatina Es Un PéptidotiterecchiutoAún no hay calificaciones
- ABSTRACTO Neuroplasticidad Aspectos BioquímicosDocumento14 páginasABSTRACTO Neuroplasticidad Aspectos BioquímicosJose PichardoAún no hay calificaciones
- Taller de Bioquímica Carbohidratos-2019Documento3 páginasTaller de Bioquímica Carbohidratos-2019Laura Tatiana Perez ForeroAún no hay calificaciones
- 2018act IV Vsa4to Biomoleculas-NvaedicionDocumento16 páginas2018act IV Vsa4to Biomoleculas-NvaedicionCarlos Javier Rivera RiveraAún no hay calificaciones
- Otros Sistemas SanguineosDocumento27 páginasOtros Sistemas SanguineosWilmer Cruzado CarrascoAún no hay calificaciones
- 2020 Proteinas en La Nutrición HumanaDocumento7 páginas2020 Proteinas en La Nutrición Humanafacundo souzaa100% (1)
- Practica de Biologia Semana 4Documento3 páginasPractica de Biologia Semana 4JANDER GONZALES ARANDAAún no hay calificaciones
- Ordinario de Biomed 2019-2Documento37 páginasOrdinario de Biomed 2019-2Leonardo Jasso PinedaAún no hay calificaciones
- AminoacidoDocumento46 páginasAminoacidoRAUL RICARDO CARHUAPOMA NICOLASAún no hay calificaciones
- Resumen Mitosis y MeiosISDocumento21 páginasResumen Mitosis y MeiosISAlgebraGeo HernandezAún no hay calificaciones
- Informe PolimorfismoDocumento15 páginasInforme PolimorfismoJuan Wilfredo Hernandez DiazAún no hay calificaciones
- Guía de Indagadores Quimica - Los LipidosDocumento5 páginasGuía de Indagadores Quimica - Los LipidosIsaac DavidAún no hay calificaciones
- Automaticidad Del OrganismoDocumento5 páginasAutomaticidad Del Organismojuan_paredes_108Aún no hay calificaciones
- CAPÍTULO 5 - Aminoácidos, Péptidos y ProteínasDocumento87 páginasCAPÍTULO 5 - Aminoácidos, Péptidos y ProteínasElliot GonzálezAún no hay calificaciones
- Pla #1 FisiologiaDocumento9 páginasPla #1 Fisiologiasamantha venturaAún no hay calificaciones
- Síntesis de PéptidosDocumento31 páginasSíntesis de PéptidosIvonne VázquezAún no hay calificaciones
- U1 - Actividad 2. ¿De Qué Están Hechas Las BiomoléculasDocumento3 páginasU1 - Actividad 2. ¿De Qué Están Hechas Las Biomoléculasstirling47% (15)
- Med S39 - "Acromegalia"Documento9 páginasMed S39 - "Acromegalia"CarlosAlbertoDangeloToledoAún no hay calificaciones
- Trabajo Sobre La Celula o Membrana CelularDocumento5 páginasTrabajo Sobre La Celula o Membrana Celularjonathan vargasAún no hay calificaciones
- TRABAJO FIN DE GRADO - HiperamonemiaDocumento20 páginasTRABAJO FIN DE GRADO - HiperamonemiaGustavo Valdelamar RuizAún no hay calificaciones
- Trabajo Escrito. Producción de InsulinaDocumento15 páginasTrabajo Escrito. Producción de InsulinaEdnaAún no hay calificaciones
- Tipificación Del ADNDocumento2 páginasTipificación Del ADNjebzsmca100% (1)
- Cuestionario Sobre Linfocitos B e InmunoglobulinasDocumento5 páginasCuestionario Sobre Linfocitos B e InmunoglobulinasDavid Tafur LeónAún no hay calificaciones
- Modelos de La Expresión GenéticaDocumento4 páginasModelos de La Expresión GenéticaGabriela CentenoAún no hay calificaciones
- Venta de Maquina de Campos Magnéticos PulsadosDocumento2 páginasVenta de Maquina de Campos Magnéticos PulsadosMago GrisAún no hay calificaciones