También podría gustarte
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Stretching: Dote a su cuerpo de una mayor flexibilidad y corrija molestas desviaciones posturalesDe EverandStretching: Dote a su cuerpo de una mayor flexibilidad y corrija molestas desviaciones posturalesCalificación: 5 de 5 estrellas5/5 (1)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Mecanismos Neuromusculares Periféricos o SensorialesDocumento12 páginasMecanismos Neuromusculares Periféricos o SensorialesSara CardeñoAún no hay calificaciones
- SLC90 ProtocoloDocumento3 páginasSLC90 ProtocoloLizbeth AraujoAún no hay calificaciones
- Tono muscular: regulación y componentes del reflejo miotáticoDocumento10 páginasTono muscular: regulación y componentes del reflejo miotáticoDaniel Aaron Fernandez GuzmanAún no hay calificaciones
- Neurofisiologia. Reflejos PDFDocumento300 páginasNeurofisiologia. Reflejos PDFCristian Moreno Madrigal100% (10)
- Bases Del Movimiento HumanoDocumento76 páginasBases Del Movimiento HumanoGrecia Melchor100% (1)
- La Propiocepción ConscienteDocumento15 páginasLa Propiocepción ConscienteMauricio100% (1)
- El Sistema Nervioso y El Movimiento 1Documento8 páginasEl Sistema Nervioso y El Movimiento 1verito canAún no hay calificaciones
- Rol Del Sistema Sensoriomotor en La Estabilidad ArticularDocumento5 páginasRol Del Sistema Sensoriomotor en La Estabilidad ArticularfasckAún no hay calificaciones
- Neurofisiologia - Alberto Gomez EstebanDocumento300 páginasNeurofisiologia - Alberto Gomez EstebanNilton RodriguezAún no hay calificaciones
- El Programa de Educación Emocional Happy 8-12Documento14 páginasEl Programa de Educación Emocional Happy 8-12Arte Pictorico Henry CubillosAún no hay calificaciones
- Sistema Motor Control Del Movimiento Reflejo y VoluntarioDocumento5 páginasSistema Motor Control Del Movimiento Reflejo y VoluntarioGonzalo Aguayo Alcayaga100% (1)
- Trabajo de Unidad 7Documento12 páginasTrabajo de Unidad 7Angel Deward Tiburcio HicianoAún no hay calificaciones
- Extra 4. El Control Del MovimientoDocumento15 páginasExtra 4. El Control Del MovimientoAlbaAún no hay calificaciones
- PROPIOCEPCIÓNDocumento6 páginasPROPIOCEPCIÓNManuel Quispe PerezAún no hay calificaciones
- TEMA 20 y 21. Control de La Actividad MotoraDocumento71 páginasTEMA 20 y 21. Control de La Actividad MotoraPaola Martin ZalukaevaAún no hay calificaciones
- Sistema Inhibidor de La Celula de RenshawDocumento8 páginasSistema Inhibidor de La Celula de RenshawMonte Sann MarielaAún no hay calificaciones
- Dialnet ElMovimientoHumano 4018449 PDFDocumento21 páginasDialnet ElMovimientoHumano 4018449 PDFLiz GuzmanAún no hay calificaciones
- tema 8 fisioDocumento23 páginastema 8 fisioa32192423Aún no hay calificaciones
- Sistemas MotoresDocumento31 páginasSistemas MotoresrominaAún no hay calificaciones
- Biomecanica ResumenDocumento28 páginasBiomecanica ResumenpaulaAún no hay calificaciones
- Trabajo (Grupo 1 Seccion 3a) Temas 19,20,21,22,23Documento35 páginasTrabajo (Grupo 1 Seccion 3a) Temas 19,20,21,22,23Elizabeth QuevedoAún no hay calificaciones
- Articulación Temporomandibular FisiologíaDocumento4 páginasArticulación Temporomandibular FisiologíaMiguel AmbrocioAún no hay calificaciones
- EJERCICIOS DE PROPIOCEPCION EN EL DEPORTEDocumento6 páginasEJERCICIOS DE PROPIOCEPCION EN EL DEPORTEcarolina.dieguez.tlqAún no hay calificaciones
- 05 Receptores Del Musculo EsqueleticoDocumento5 páginas05 Receptores Del Musculo EsqueleticoMayka RamosAún no hay calificaciones
- Seminario Grupo 3-1Documento20 páginasSeminario Grupo 3-1rogelys palenciaAún no hay calificaciones
- Unidad 3 de FisiologiaDocumento5 páginasUnidad 3 de FisiologiaMalena Torres PozoAún no hay calificaciones
- MotilidadDocumento49 páginasMotilidadTadeo RamosAún no hay calificaciones
- Órgano Tendinoso de GolgiDocumento5 páginasÓrgano Tendinoso de GolgiYesell Betsabeth Pulido ValeroAún no hay calificaciones
- Los Sistemas Efectores - Psicología UNEDDocumento8 páginasLos Sistemas Efectores - Psicología UNEDsugeypaezAún no hay calificaciones
- Adf MasticacionDocumento33 páginasAdf MasticacionESTEFANIA MATTOS MARINAún no hay calificaciones
- Bases neurofisiológicas de la propiocepciónDocumento38 páginasBases neurofisiológicas de la propiocepciónAntonio Sanchez100% (1)
- Sensibilidad PropioceptivaDocumento10 páginasSensibilidad PropioceptivaMaría Sanchez GAún no hay calificaciones
- Reflejo MedularDocumento5 páginasReflejo MedularMaily MuñozAún no hay calificaciones
- PROPIOCEPCIÓNDocumento14 páginasPROPIOCEPCIÓNgrabiel IrigarayAún no hay calificaciones
- Certamen 2 - FisioDocumento52 páginasCertamen 2 - Fisioisidora zellerAún no hay calificaciones
- Sistema MotorDocumento7 páginasSistema MotorRamírez OmarAún no hay calificaciones
- Entrenamiento FuncionalDocumento26 páginasEntrenamiento FuncionalMiguel MurgaAún no hay calificaciones
- Sistema PropioceptivoDocumento6 páginasSistema PropioceptivoKalu KalucashAún no hay calificaciones
- Tema 7Documento10 páginasTema 7paula clua torresAún no hay calificaciones
- 10 Sistema Neuromuscular Documento PDFDocumento22 páginas10 Sistema Neuromuscular Documento PDFpablomyloveAún no hay calificaciones
- Cuestiones Fisiología MuscularDocumento7 páginasCuestiones Fisiología Muscularmaria del carmen ruiz fernandezAún no hay calificaciones
- Capítulo 8 Control Del Movimiento. ResumenDocumento15 páginasCapítulo 8 Control Del Movimiento. Resumenmilagrosperalta5382Aún no hay calificaciones
- Organizacion MotricidadDocumento6 páginasOrganizacion MotricidadECAAún no hay calificaciones
- Sensibilidad PropioceptivaDocumento22 páginasSensibilidad PropioceptivaMichael DuranAún no hay calificaciones
- Reflejo Medular: ClasificaciónDocumento6 páginasReflejo Medular: ClasificaciónJoab Lira DiazAún no hay calificaciones
- 3.2.sensibilidad PropioceptivaDocumento12 páginas3.2.sensibilidad PropioceptivaDony ZamoranoAún no hay calificaciones
- Sistema nervioso y coordinación motora en yogaDocumento8 páginasSistema nervioso y coordinación motora en yogaYoga FlowAún no hay calificaciones
- Funcion NeuromuscularDocumento13 páginasFuncion NeuromuscularAdriana Cardeña100% (1)
- Regulación MuscularDocumento12 páginasRegulación MuscularJoseRvsAún no hay calificaciones
- MÚSCULOSDocumento5 páginasMÚSCULOSCecilia Murrieta LimonAún no hay calificaciones
- Huso NeuromuscularDocumento33 páginasHuso NeuromuscularGab GahanAún no hay calificaciones
- 2do Informe TerminadoDocumento38 páginas2do Informe TerminadoEdwin Antonio Garcia VasquezAún no hay calificaciones
- Órganos Terminales PropioceptivosDocumento8 páginasÓrganos Terminales PropioceptivosDavid BarrazaAún no hay calificaciones
- Sensibilidad PropioceptivaDocumento11 páginasSensibilidad PropioceptivaKarla ArriagadaAún no hay calificaciones
- Funciones Motoras de La Medula Espinal Reflejos Medulares Cap 54Documento19 páginasFunciones Motoras de La Medula Espinal Reflejos Medulares Cap 54Abraham Alvarado0% (2)
- Hatesse Tecnica Tendinosa PDFDocumento6 páginasHatesse Tecnica Tendinosa PDFJosè Manuel Pinto LopezAún no hay calificaciones
- A Flor de Piel - Lena ValentiDocumento243 páginasA Flor de Piel - Lena Valentilic.tatiana.k.perezAún no hay calificaciones
- Examen GCHDocumento31 páginasExamen GCHSIR EDISONAún no hay calificaciones
- Empoderamiento - David Aguado. Profesor de La UAMDocumento7 páginasEmpoderamiento - David Aguado. Profesor de La UAMPablo Alonso ArduraAún no hay calificaciones
- Teorias de La Motivacin. Aplicacin PrcticaDocumento108 páginasTeorias de La Motivacin. Aplicacin PrcticaDulce Belem Canul CuellarAún no hay calificaciones
- Lectura de Casos - Estrategias en AulaDocumento35 páginasLectura de Casos - Estrategias en AulaCdph Mentes GenialesAún no hay calificaciones
- Javier AliagaDocumento308 páginasJavier AliagaWilber CallisayaAún no hay calificaciones
- 05 Detección de Maltrato en La Persona MayorDocumento1 página05 Detección de Maltrato en La Persona MayorDalia RodriguezAún no hay calificaciones
- Leahey Habla de 4 Formas de Entender El MundoDocumento2 páginasLeahey Habla de 4 Formas de Entender El MundoIvana MolinaAún no hay calificaciones
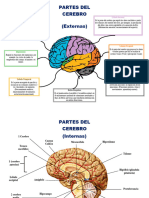
- Partes Del CerebroDocumento2 páginasPartes Del CerebroNAYELI GARCIAS RESENDIZAún no hay calificaciones
- Unidad 2Documento15 páginasUnidad 2Michelle DominguezAún no hay calificaciones
- CognitivismoDocumento18 páginasCognitivismopaolamontalvancAún no hay calificaciones
- Taller Etica 3Documento7 páginasTaller Etica 3Diego Fernando Ruiz ZapataAún no hay calificaciones
- Planificación de Una Clase para El 28 - 09 - 2022Documento5 páginasPlanificación de Una Clase para El 28 - 09 - 2022Mariana CardozoAún no hay calificaciones
- GUIA DE REPASO DE CIENCIAS 4°cuerpohumanoDocumento3 páginasGUIA DE REPASO DE CIENCIAS 4°cuerpohumanocoral herrera carrasco100% (1)
- Evaluacion Juego Migdalia..Documento2 páginasEvaluacion Juego Migdalia..karla vinuezaAún no hay calificaciones
- Caso ChocburysDocumento3 páginasCaso ChocburysRODRIGO BENJAMIN GUTIERREZ GANDULFOAún no hay calificaciones
- Tarea 1 P. DesarrolloDocumento12 páginasTarea 1 P. DesarrolloLaura RosarioAún no hay calificaciones
- Educación y tecnología clave para el futuroDocumento4 páginasEducación y tecnología clave para el futuroTatiana FrancoAún no hay calificaciones
- 1SESIONDocumento32 páginas1SESIONjuanita100% (1)
- Hoja de Respuestas. GATES.Documento3 páginasHoja de Respuestas. GATES.gofebi7537Aún no hay calificaciones
- Sesion 1. SolidaridadDocumento12 páginasSesion 1. SolidaridadGRAL.MARIANO ESCOBEDO 571JAún no hay calificaciones
- Conceptos Básicos de SexualidadDocumento4 páginasConceptos Básicos de SexualidadDylan CollinsAún no hay calificaciones
- Prueba Sistema Nervioso Cuarto BásicoDocumento4 páginasPrueba Sistema Nervioso Cuarto BásicoJess SiCaAún no hay calificaciones
- 3 - Miller - El Inconsciente y El Cuerpo HablanteDocumento7 páginas3 - Miller - El Inconsciente y El Cuerpo HablanteLauraAún no hay calificaciones
- Practica 1 Metodo CientificoDocumento10 páginasPractica 1 Metodo CientificomileAún no hay calificaciones
- Etapa Del Desarrollo.Documento1 páginaEtapa Del Desarrollo.JUAN PABLO RENGIFOAún no hay calificaciones
- Análisis fenomenológico de las diferencias entre la vista y el tactoDocumento3 páginasAnálisis fenomenológico de las diferencias entre la vista y el tactoElisa LamasAún no hay calificaciones
- Sistemas Duros y BlandoDocumento3 páginasSistemas Duros y BlandoHenry Pinedo ArizaAún no hay calificaciones