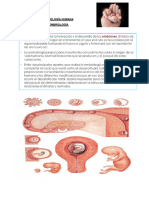
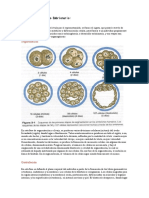
También podría gustarte
- La GastrulaciónDocumento12 páginasLa GastrulaciónSandra Jakelin Peralta CotrinaAún no hay calificaciones
- Lectura - Gastrulación, Proceso Clave en La Formación de Un Nuevo OrganismoDocumento15 páginasLectura - Gastrulación, Proceso Clave en La Formación de Un Nuevo OrganismoPEDRO MANUEL CASTRO RIVERAAún no hay calificaciones
- Gastrulación: Proceso clave en la formación de capas germinalesDocumento11 páginasGastrulación: Proceso clave en la formación de capas germinalesEden Mata RAún no hay calificaciones
- Desarrollo EmbrionarioDocumento12 páginasDesarrollo EmbrionarioMarilyn Kilzi100% (1)
- Embriología Tarea 1Documento18 páginasEmbriología Tarea 1camila moranAún no hay calificaciones
- Formación Disco Germinativo TrilaminarDocumento18 páginasFormación Disco Germinativo TrilaminarSebastian AmpueroAún no hay calificaciones
- histoligi 4Documento14 páginashistoligi 4dfAún no hay calificaciones
- Reporte de Lab Embriogénesis TempranaDocumento10 páginasReporte de Lab Embriogénesis Tempranacv22036Aún no hay calificaciones
- Embrión TrilaminarDocumento3 páginasEmbrión TrilaminarLAURA MARCELA BARRENECHE CALLEAún no hay calificaciones
- Cuestionario II EmbriologiaDocumento11 páginasCuestionario II EmbriologiaAnahis CardenasAún no hay calificaciones
- Tipos de FecundidadDocumento10 páginasTipos de FecundidadBetanzos Estrada RalfherAún no hay calificaciones
- Desarrollo EmbrionarioDocumento10 páginasDesarrollo EmbrionarioPJAún no hay calificaciones
- Trabajo ShereDocumento11 páginasTrabajo ShereMini moto 46Aún no hay calificaciones
- Fecundaciontema 3Documento13 páginasFecundaciontema 3Vilma RamirezAún no hay calificaciones
- Desarrollo EmbrionarioDocumento10 páginasDesarrollo EmbrionarioJesus Enrique Peña CoronadoAún no hay calificaciones
- Etapas Del Desarrollo 1Documento5 páginasEtapas Del Desarrollo 1c7gdp8mkyfAún no hay calificaciones
- SEGMENTACIONDocumento11 páginasSEGMENTACIONrosmeryzambrano05Aún no hay calificaciones
- Teorías del desarrollo embrionarioDocumento7 páginasTeorías del desarrollo embrionarioageninazarAún no hay calificaciones
- EMBRIOLOGÍA HUMANADocumento10 páginasEMBRIOLOGÍA HUMANAMaría Angélica SosaAún no hay calificaciones
- Guía sobre embriología y gametogénesisDocumento7 páginasGuía sobre embriología y gametogénesisMabelAún no hay calificaciones
- GUIA 1 Desarrollo Embrionario 2020 alumnosUTDocumento8 páginasGUIA 1 Desarrollo Embrionario 2020 alumnosUTMARIA CAMILA PINA ESPINOSAAún no hay calificaciones
- Lateralidad EmbriologiaDocumento5 páginasLateralidad EmbriologiaJeronimo RosasAún no hay calificaciones
- Gestación en AnimalesDocumento23 páginasGestación en AnimalesgeraaAún no hay calificaciones
- Desarrollo Embrionario de Las AvesDocumento15 páginasDesarrollo Embrionario de Las AvesAna Mendoza0% (2)
- Práctica 8 (Desarrollo Embrionario) Parte 2 Grupo 1Documento9 páginasPráctica 8 (Desarrollo Embrionario) Parte 2 Grupo 1Yankuam AntunAún no hay calificaciones
- Trabajo de BiologiaDocumento37 páginasTrabajo de BiologiaYakelin CastellanoAún no hay calificaciones
- Tema EmbriologiaDocumento18 páginasTema EmbriologiaSofia SirrAún no hay calificaciones
- Desarrollo embrionario: etapas de la fecundación, segmentación, gastrulación y organogénesisDocumento8 páginasDesarrollo embrionario: etapas de la fecundación, segmentación, gastrulación y organogénesismarialejandraAún no hay calificaciones
- Preparación del aparato reproductor femenino para la gestaciónDocumento13 páginasPreparación del aparato reproductor femenino para la gestaciónGabriela de LeónAún no hay calificaciones
- Tema 1Documento15 páginasTema 1luciamartinizquierdoAún no hay calificaciones
- Ensayo EmbriologiaDocumento6 páginasEnsayo EmbriologiajohanaAún no hay calificaciones
- Origen y desarrollo embrionarioDocumento6 páginasOrigen y desarrollo embrionarioMaria Alejandra Zamora LopezAún no hay calificaciones
- Informe sobre las etapas del desarrollo embrionarioDocumento8 páginasInforme sobre las etapas del desarrollo embrionariopayalag232Aún no hay calificaciones
- Zonas de Referencia y Planos de Corte Del EmbriónDocumento11 páginasZonas de Referencia y Planos de Corte Del EmbriónArancibia Leon Giselle AydeeAún no hay calificaciones
- El Desarrollo EmbrionarioDocumento3 páginasEl Desarrollo EmbrionarioFrank CeballosAún no hay calificaciones
- Definición de CélulaDocumento2 páginasDefinición de Célulajenipino83Aún no hay calificaciones
- Fases Del Desarrollo EmbrionarioDocumento2 páginasFases Del Desarrollo Embrionarioclara riveiroAún no hay calificaciones
- 4TA A 8VA SEMANA A PDFDocumento12 páginas4TA A 8VA SEMANA A PDFMaryori Treyci ILasaca ValenzuelaAún no hay calificaciones
- Desarrollo embrionario porcinoDocumento5 páginasDesarrollo embrionario porcinoJhojan Victorio ChachiAún no hay calificaciones
- Tarea 4 ReproDocumento6 páginasTarea 4 Repromayra martinezAún no hay calificaciones
- ApuntesembriologiaDocumento131 páginasApuntesembriologiasylarhillsAún no hay calificaciones
- Desarrollo Embrionario y Fetal HumanoDocumento4 páginasDesarrollo Embrionario y Fetal HumanoMîîrlêîîvîîsThûÔdîîôxîîthâAún no hay calificaciones
- Primera Semana Del Desarrollo EmbrionarioDocumento8 páginasPrimera Semana Del Desarrollo Embrionariojuan pabloAún no hay calificaciones
- Gastrulación y desarrollo embrionario: formación de capas celulares y órganosDocumento4 páginasGastrulación y desarrollo embrionario: formación de capas celulares y órganosandres ramosAún no hay calificaciones
- PlacentacionDocumento23 páginasPlacentacionHugo Quiles DetweilerAún no hay calificaciones
- EMBRIOLOGIA ResumenDocumento39 páginasEMBRIOLOGIA ResumenFran Madriaza50% (4)
- Embriología Primeras SemanasDocumento16 páginasEmbriología Primeras SemanasHamilton AndradeAún no hay calificaciones
- Guía para El Estudio de La EmbriologíaDocumento14 páginasGuía para El Estudio de La EmbriologíaMaria CejasAún no hay calificaciones
- Embriologia Clase 1Documento9 páginasEmbriologia Clase 1DELIAAún no hay calificaciones
- Segmentación embrionaria y gastrulación en la primera semanaDocumento7 páginasSegmentación embrionaria y gastrulación en la primera semanaYessii AlonzoAún no hay calificaciones
- Reproduccion CelularDocumento27 páginasReproduccion CelularVicky Jimenez100% (1)
- Embriología Humana General.Documento9 páginasEmbriología Humana General.olenka marinAún no hay calificaciones
- Nociones de EmbriologiaDocumento11 páginasNociones de EmbriologiaOriana Hnnd'z-m0% (1)
- BLASTOGÉNESISDocumento5 páginasBLASTOGÉNESISImelda Moreno100% (1)
- Trabajo 2 BiologiaDocumento18 páginasTrabajo 2 BiologiaDiego MartínezAún no hay calificaciones
- Wa0141.Documento26 páginasWa0141.VictoriaAún no hay calificaciones
- Guía Desarrollo EmbrionarioDocumento10 páginasGuía Desarrollo EmbrionarioHéctor Morales Ponce0% (1)
- Desarrollo embrionario tempranoDocumento20 páginasDesarrollo embrionario tempranoAlex Port0% (1)
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSDe EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- Fibromialgia y OsteopatiaDocumento42 páginasFibromialgia y OsteopatiaMirtaAún no hay calificaciones
- Condroproteccion 47248903Documento34 páginasCondroproteccion 47248903MirtaAún no hay calificaciones
- SNA Anual 2017Documento172 páginasSNA Anual 2017Cátedra de Farmacología de la Escuela de Medicina de La UNIVERSIDAD DEL ZULIA100% (6)
- Anatomía de La FasciaDocumento4 páginasAnatomía de La FasciaMirtaAún no hay calificaciones
- Herbert Spencer y sus Primeros PrincipiosDocumento291 páginasHerbert Spencer y sus Primeros PrincipiosjlmansillaAún no hay calificaciones
- eMOCIONES EN MOVIMIENTO-INTEROCEPCION FASCIAL - ESPAÑOLDocumento6 páginaseMOCIONES EN MOVIMIENTO-INTEROCEPCION FASCIAL - ESPAÑOLMirtaAún no hay calificaciones
- La Exploracion Del Torax Una Guia para DDocumento15 páginasLa Exploracion Del Torax Una Guia para DMirtaAún no hay calificaciones
- Coronavirus Natura EbookDocumento184 páginasCoronavirus Natura EbookMirtaAún no hay calificaciones
- OSTEO PsicosomaticaDocumento14 páginasOSTEO PsicosomaticaMirta100% (1)
- Neu To Transm I SoresDocumento24 páginasNeu To Transm I SoresLeonardo GiraldoAún no hay calificaciones
- Linfaticos de Cuello y Organos de Los SentidosDocumento17 páginasLinfaticos de Cuello y Organos de Los SentidosMirtaAún no hay calificaciones
- Texto Español-Fluido LCR y Linfa BCH (Copia Conflictiva de LUCI HERRERA 2012-06-18)Documento13 páginasTexto Español-Fluido LCR y Linfa BCH (Copia Conflictiva de LUCI HERRERA 2012-06-18)MirtaAún no hay calificaciones
- Emocional y OsteopatíaDocumento12 páginasEmocional y OsteopatíaMirtaAún no hay calificaciones
- Impact of Chronic Stress On Individual HDocumento13 páginasImpact of Chronic Stress On Individual HMirtaAún no hay calificaciones
- Integración de Señales Gastrointestinales Vestibulares y Eméticas Que Producir Náuseas y VómitosDocumento31 páginasIntegración de Señales Gastrointestinales Vestibulares y Eméticas Que Producir Náuseas y VómitosMirtaAún no hay calificaciones
- Reglas para Manipulacion Vertebral Jy MaigneDocumento13 páginasReglas para Manipulacion Vertebral Jy MaigneAntonioAún no hay calificaciones
- Johns Hopkins Medicine International LaDocumento30 páginasJohns Hopkins Medicine International LaMirtaAún no hay calificaciones
- Efectos Del Tratamiento Del Musculo DiafDocumento267 páginasEfectos Del Tratamiento Del Musculo DiafMirtaAún no hay calificaciones
- Base Scientific As Art Ritis Reuma To I deDocumento22 páginasBase Scientific As Art Ritis Reuma To I deDiana Cecilia Cabana OrozcoAún no hay calificaciones
- Neuroanatomia Funcional AfifiDocumento451 páginasNeuroanatomia Funcional Afififogony92% (153)
- Tronco Interno y ExternoDocumento32 páginasTronco Interno y ExternoMirtaAún no hay calificaciones
- MarthaCeciliaOspinaGuisao 2019Documento60 páginasMarthaCeciliaOspinaGuisao 2019Pato LezcanoAún no hay calificaciones
- El Tejido Óseo Es Una Parte Integral Del Sistema FascialDocumento3 páginasEl Tejido Óseo Es Una Parte Integral Del Sistema FascialMirtaAún no hay calificaciones
- Anatomía de La Fascia AbdominalDocumento4 páginasAnatomía de La Fascia AbdominalMirtaAún no hay calificaciones
- El Otro Lado de La Fascia PARTE 2Documento6 páginasEl Otro Lado de La Fascia PARTE 2MirtaAún no hay calificaciones
- Relación anatómica de los ganglios simpáticos cervicales y celiacos con la terapia neuralDocumento98 páginasRelación anatómica de los ganglios simpáticos cervicales y celiacos con la terapia neuralCamilo100% (2)
- Caracterización Morfológica y Molecular de Colectores Linfáticos Dérmicos HumanosDocumento14 páginasCaracterización Morfológica y Molecular de Colectores Linfáticos Dérmicos HumanosMirtaAún no hay calificaciones
- Suelo PelvicoDocumento68 páginasSuelo PelvicoZulma QuispeAún no hay calificaciones
- Tendon Valoracion y Tto FisioterapiaDocumento311 páginasTendon Valoracion y Tto FisioterapiaValentina Macarena Diaz Jara89% (18)
- Efectos Del Tratamiento Del Musculo DiafDocumento267 páginasEfectos Del Tratamiento Del Musculo DiafMirtaAún no hay calificaciones
- Utpl Tnbif001g PDFDocumento118 páginasUtpl Tnbif001g PDFAnonymous GCe6aeGzL100% (1)
- HerbicidasDocumento2 páginasHerbicidasGabo Bernal OcegueraAún no hay calificaciones
- Tumores de Tórax TACDocumento14 páginasTumores de Tórax TACArantza SalazarAún no hay calificaciones
- Anatomía de Los VentrículosDocumento2 páginasAnatomía de Los Ventrículosjosselyn.hdez01Aún no hay calificaciones
- Bases para DentadurasDocumento4 páginasBases para DentadurasKaren GarciaAún no hay calificaciones
- Crisis HTADocumento9 páginasCrisis HTACLAUDIA JAVIERA ANTUNEZ LAYAún no hay calificaciones
- Sobredentaduras sobre perno tapaDocumento4 páginasSobredentaduras sobre perno tapaPaula Morales100% (5)
- Trabajo Del Sistema Cardio VascularDocumento14 páginasTrabajo Del Sistema Cardio Vascularjesús jáureguiAún no hay calificaciones
- Desórdenes Motores Del Habla y PROMPTDocumento9 páginasDesórdenes Motores Del Habla y PROMPTSamy SanAún no hay calificaciones
- Senescencia Lesiones de AlmacenamientoDocumento6 páginasSenescencia Lesiones de Almacenamientocatalina AlcarazAún no hay calificaciones
- Semiologia de Abdomen....Documento27 páginasSemiologia de Abdomen....Mariela Isabell Ceballos100% (1)
- Bioseguridad y Manejo de CitostaticosDocumento3 páginasBioseguridad y Manejo de CitostaticosKimberly Melissa MadridAún no hay calificaciones
- Los sustantivos y sus característicasDocumento102 páginasLos sustantivos y sus característicasvictor425Aún no hay calificaciones
- Curso de Canto en 10 PasosDocumento7 páginasCurso de Canto en 10 PasosIsaac PeretzAún no hay calificaciones
- Inervación y Vascularización de La Cara y La Piel Cabelluda.Documento4 páginasInervación y Vascularización de La Cara y La Piel Cabelluda.EdithAún no hay calificaciones
- 10) Factores de Virulencia Bacterianos y PatogenicidadDocumento12 páginas10) Factores de Virulencia Bacterianos y PatogenicidadZósimo SuasnabarAún no hay calificaciones
- Diarrea Aguda y CrónicaDocumento33 páginasDiarrea Aguda y CrónicaGaby FernandaAún no hay calificaciones
- Grupo 3 Expo # 02 (Hueso de La Cara)Documento25 páginasGrupo 3 Expo # 02 (Hueso de La Cara)Guillermo Macharé UbillúsAún no hay calificaciones
- DiureticosDocumento53 páginasDiureticosMiguel BordaAún no hay calificaciones
- Farmacodinamia: Receptores, Agonistas y AntagonistasDocumento10 páginasFarmacodinamia: Receptores, Agonistas y AntagonistasJosé Gustavo Gaitán FloresAún no hay calificaciones
- Vasos Maravillosos Ancestrales Acupuntura y Medicina Natural ArequipaDocumento15 páginasVasos Maravillosos Ancestrales Acupuntura y Medicina Natural Arequipashybumi100% (1)
- Regulación del calcio y fósforo séricoDocumento63 páginasRegulación del calcio y fósforo séricoAndres RestrepoAún no hay calificaciones
- Los Musculos 2Documento51 páginasLos Musculos 2Dionisio FVAún no hay calificaciones
- Postura corporal guíaDocumento6 páginasPostura corporal guíaMarcos AntonioAún no hay calificaciones
- Anatomia Forense 1225582479937259 8Documento25 páginasAnatomia Forense 1225582479937259 8Compañerin CixAún no hay calificaciones
- CitologíaDocumento48 páginasCitologíaasancheztome1550Aún no hay calificaciones
- La estructura y funciones de la célulaDocumento11 páginasLa estructura y funciones de la célulaJAIRO DE JESUS LASCARRO ESCORCIAAún no hay calificaciones
- Consolidacion Pato I Tema 3Documento7 páginasConsolidacion Pato I Tema 3abdallah hamdanAún no hay calificaciones
- Libro Veterinaria Practica 2018 Pediatria Chile PDFDocumento223 páginasLibro Veterinaria Practica 2018 Pediatria Chile PDFCarolina Andrea Cerda ParedesAún no hay calificaciones
- Sesión 8Documento6 páginasSesión 8Orlando TrejoAún no hay calificaciones