También podría gustarte
- Examen de La Mujer GrávidaDocumento21 páginasExamen de La Mujer GrávidaAntonella GómezAún no hay calificaciones
- Casos Clínicos 1Documento4 páginasCasos Clínicos 1erika moralesAún no hay calificaciones
- Radiografía de tórax: proyecciones y parámetros normalesDocumento42 páginasRadiografía de tórax: proyecciones y parámetros normalesGabriela VillalobosAún no hay calificaciones
- Minimanual Ginecologia - CTO Editado PDFDocumento28 páginasMinimanual Ginecologia - CTO Editado PDFariesyaraAún no hay calificaciones
- Posiciones Del ÚteroDocumento1 páginaPosiciones Del ÚteroCarmen Esquivel ZarzaAún no hay calificaciones
- Resume OrlDocumento21 páginasResume OrlRodrigo Andres Vega DiazAún no hay calificaciones
- Trauma VascularDocumento6 páginasTrauma VascularMAI100% (1)
- Fibrilacion AuricularDocumento41 páginasFibrilacion AuricularyinfsAún no hay calificaciones
- Escroto Agudo en NiñosDocumento34 páginasEscroto Agudo en Niñosrccm_1010Aún no hay calificaciones
- Neumonía Adquirida en La Comunidad PDFDocumento18 páginasNeumonía Adquirida en La Comunidad PDFLinda MedinaAún no hay calificaciones
- Pancreatitis 2020Documento35 páginasPancreatitis 2020Yury Rojas SaenzAún no hay calificaciones
- Medicina LegalDocumento11 páginasMedicina LegalSare YoisAún no hay calificaciones
- Cáncer de Ampolla de VaterDocumento6 páginasCáncer de Ampolla de VaterHelfer Fernando Aquino Pilco100% (1)
- Examen de La Mujer GravidaDocumento15 páginasExamen de La Mujer Gravidagarfield_12Aún no hay calificaciones
- Aparato reproductor masculino: trastornos y enfermedadesDocumento25 páginasAparato reproductor masculino: trastornos y enfermedadesMarielos RodriguezAún no hay calificaciones
- 20a Sesión SNyE Funciones de Las Gónadas Masc y FemDocumento52 páginas20a Sesión SNyE Funciones de Las Gónadas Masc y FemRuth Cobo RosalesAún no hay calificaciones
- Dolor abdominal pediatría causas diagnósticoDocumento21 páginasDolor abdominal pediatría causas diagnósticoCatherine Andrea Olate SaavedraAún no hay calificaciones
- Neuro CirDocumento60 páginasNeuro CirZeptAlanAún no hay calificaciones
- Luxaciones de Hombro y CaderaDocumento28 páginasLuxaciones de Hombro y CaderaMario Andres Flores100% (1)
- Politraumatismo en PediatriaDocumento53 páginasPolitraumatismo en PediatriaXimena MaitaAún no hay calificaciones
- Dolor abdominal agudo: causas y tratamientoDocumento35 páginasDolor abdominal agudo: causas y tratamientoJose Luis Raymundo Perea100% (1)
- Ultrafiltracion renal y hemodinamia glomerularDocumento7 páginasUltrafiltracion renal y hemodinamia glomerularArturo Oviedo Chávez100% (2)
- Resumen GPC AsmaDocumento12 páginasResumen GPC AsmaeherediaaAún no hay calificaciones
- Clase Infectologia IDocumento173 páginasClase Infectologia IAlmeyda JeffersonAún no hay calificaciones
- RadioinmunoensayoDocumento13 páginasRadioinmunoensayoRicardo Perez EspadaAún no hay calificaciones
- Exploración de AbdomenDocumento22 páginasExploración de AbdomenYayi Magaña0% (1)
- Oftalmología MIR 2004/2005: Enfermedades orbitarias y externasDocumento17 páginasOftalmología MIR 2004/2005: Enfermedades orbitarias y externasANGUIE ANETTE MAMANI SUCASAIREAún no hay calificaciones
- Sepsis PDFDocumento11 páginasSepsis PDFMiguel Vasquez Sanchez0% (1)
- Enfermedad de CrohnDocumento41 páginasEnfermedad de CrohnRafa RodriguezAún no hay calificaciones
- Cancer de Mama Final Re Contra FinalDocumento37 páginasCancer de Mama Final Re Contra FinalValeria Heredia Peña100% (1)
- Atlas de Diagnóstico y Tratamiento en Neumología PDFDocumento26 páginasAtlas de Diagnóstico y Tratamiento en Neumología PDFAngel GamboaAún no hay calificaciones
- Esquema ObstetriciaDocumento23 páginasEsquema ObstetriciaSergio PerezAún no hay calificaciones
- Taller Casos Clinicos - PolitraumaDocumento13 páginasTaller Casos Clinicos - PolitraumaCristhian Penagos ParraAún no hay calificaciones
- TOXOPLASMOSISDocumento50 páginasTOXOPLASMOSISJessica Fernanda Alvarado Yumisaca100% (2)
- Doppler en screening de I trimestre y situaciones clínicasDocumento68 páginasDoppler en screening de I trimestre y situaciones clínicasfelix camposAún no hay calificaciones
- Historia Clinica PediatriaDocumento5 páginasHistoria Clinica PediatriaNatalia AquinoAún no hay calificaciones
- Perlas NeurologiaDocumento28 páginasPerlas NeurologiaDana C. ArmengolAún no hay calificaciones
- Libro Oftalmologia en Atencion PrimariaDocumento8 páginasLibro Oftalmologia en Atencion PrimariaGleison AlvesAún no hay calificaciones
- CabreritaDocumento23 páginasCabreritaRicardo Ponce SilvaAún no hay calificaciones
- Libro Del Est - CardDocumento56 páginasLibro Del Est - CardPedro Jose LopezAún no hay calificaciones
- Perforacion Viscera HuecaDocumento2 páginasPerforacion Viscera Huecafruben19Aún no hay calificaciones
- MECANISMOS INMUNOLÓGICOS Y CELULARES DE LA LESIÓN GLOMERULARDocumento13 páginasMECANISMOS INMUNOLÓGICOS Y CELULARES DE LA LESIÓN GLOMERULARChristian Itandehui JiménezAún no hay calificaciones
- Apuntes de CirugiaDocumento7 páginasApuntes de Cirugiapaola llamocuroAún no hay calificaciones
- Ley Federal Del Trabajo y Su Relacion Medico LegalDocumento35 páginasLey Federal Del Trabajo y Su Relacion Medico LegalAristides H Garcia ReyesAún no hay calificaciones
- Malabsorción y maldigestiónDocumento20 páginasMalabsorción y maldigestiónAbelAún no hay calificaciones
- 1 Ra Oftalmologia Se 387759 Downloadable 152751Documento27 páginas1 Ra Oftalmologia Se 387759 Downloadable 152751ingridupsjb100% (1)
- Trastornos motores esofágicosDocumento5 páginasTrastornos motores esofágicosPablo BosaAún no hay calificaciones
- Perdida Gestacional Primer TrimestreDocumento22 páginasPerdida Gestacional Primer TrimestreLuisa OrozcoAún no hay calificaciones
- Cuál Es La Medición Ecográfica Que Posee Durante Todo El Embarazo Un Crecimiento Lineal y ConstanteDocumento8 páginasCuál Es La Medición Ecográfica Que Posee Durante Todo El Embarazo Un Crecimiento Lineal y ConstanteJorge Alberto HernandezAún no hay calificaciones
- HEMATOLOGIA CtoDocumento25 páginasHEMATOLOGIA CtoIvalicia Ramos FujarteAún no hay calificaciones
- Historia Clinica GinecologicaDocumento11 páginasHistoria Clinica GinecologicaBryan Yucra GarciaAún no hay calificaciones
- SOPQ: Síndrome de Ovarios PoliquísticosDocumento48 páginasSOPQ: Síndrome de Ovarios PoliquísticosGrecia Torres ValderramaAún no hay calificaciones
- Solución de cloruro de sodio 10% para reposición de electrolitosDocumento12 páginasSolución de cloruro de sodio 10% para reposición de electrolitosVanessa Rocha MolinaAún no hay calificaciones
- Espina BifidaDocumento61 páginasEspina BifidaCarla OlivosAún no hay calificaciones
- Hipertensión Pulmonar Arterial: (Ascitis) En Pollos De EngordaDe EverandHipertensión Pulmonar Arterial: (Ascitis) En Pollos De EngordaAún no hay calificaciones
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleDe EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleAún no hay calificaciones
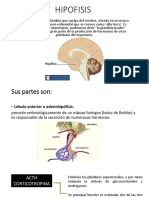
- HIPOFISISDocumento10 páginasHIPOFISISFrancis NicoleAún no hay calificaciones
- Estructura y función del aparato reproductor femeninoDocumento55 páginasEstructura y función del aparato reproductor femeninoJulissa VizaAún no hay calificaciones
- De00004 TrabajodetitulacionDocumento68 páginasDe00004 TrabajodetitulacionCOSTO OMARSAAún no hay calificaciones
- Control de La Secrecion HormonalDocumento5 páginasControl de La Secrecion HormonalGeraldyne HernandezAún no hay calificaciones
- Sangrado Uterino Anormal en La PerimenopausiaDocumento11 páginasSangrado Uterino Anormal en La PerimenopausiaValeria CallapaAún no hay calificaciones
- Cuestic3b3n de HuevosDocumento6 páginasCuestic3b3n de HuevosDaniloAún no hay calificaciones
- Anatomia y Fisiologia de La Reproduccion BovinaDocumento6 páginasAnatomia y Fisiologia de La Reproduccion Bovinachristian quinterosAún no hay calificaciones
- Embriologia Usmp SeminarioDocumento9 páginasEmbriologia Usmp SeminarioLuz Valderrama DurandAún no hay calificaciones
- Paso 1 Contextualización de Anatomía y Fisiología ReproductivaDocumento9 páginasPaso 1 Contextualización de Anatomía y Fisiología ReproductivaEsteban Fino100% (1)
- Preguntas Sistema Reproductor MasculinoDocumento4 páginasPreguntas Sistema Reproductor MasculinoCatalina Andrea Soto100% (5)
- Reproducción Sexual en AnimalesDocumento25 páginasReproducción Sexual en AnimalesSteve Vladimir Acedo LazoAún no hay calificaciones
- Causas de folículos anovulatorios en yeguas y su relación con hallazgos ecográficosDocumento38 páginasCausas de folículos anovulatorios en yeguas y su relación con hallazgos ecográficosJazmin GutiérrezAún no hay calificaciones
- B.II 2do ParcialDocumento10 páginasB.II 2do ParcialRUTH KAREN SANCHEZ LEDEZMAAún no hay calificaciones
- Fisiologia Del Eje H.H.O.E.Documento36 páginasFisiologia Del Eje H.H.O.E.jhonantanaguiarAún no hay calificaciones
- Ciclo MenstrualDocumento33 páginasCiclo MenstrualFelipeMirandaAún no hay calificaciones
- Tesis-Cuyes ModificadoDocumento26 páginasTesis-Cuyes ModificadoGiovani AltezAún no hay calificaciones
- FemeninoDocumento7 páginasFemeninoVato TorresAún no hay calificaciones
- Tesis Inseminacion Artificial PDFDocumento85 páginasTesis Inseminacion Artificial PDFManuel FloresAún no hay calificaciones
- Reproducción en camélidos sudamericanosDocumento38 páginasReproducción en camélidos sudamericanosJhonatan Juno VargasAún no hay calificaciones
- Ppa HormonasDocumento29 páginasPpa HormonasLizbeth HuertaAún no hay calificaciones
- Guía - Primer Parcial - EmbriologíaDocumento89 páginasGuía - Primer Parcial - EmbriologíaDaniela Figueroa100% (1)
- Síntomas y Duración Del Puerperio, Manejo de La Yegua en Celo y Manifestaciones Externas FotoperiodoDocumento8 páginasSíntomas y Duración Del Puerperio, Manejo de La Yegua en Celo y Manifestaciones Externas FotoperiodoFrancisco AguilarAún no hay calificaciones
- 2021.05.12 ExtraOrd B Admision 2021 IIDocumento26 páginas2021.05.12 ExtraOrd B Admision 2021 IIPlanner MantenimientoAún no hay calificaciones
- TP 2 Biologia B ResueltoDocumento28 páginasTP 2 Biologia B ResueltoClaudia NuñezAún no hay calificaciones
- 4 TiroidesDocumento103 páginas4 TiroidesChris VitaleAún no hay calificaciones
- UntitledDocumento19 páginasUntitledHector Serrano PulidoAún no hay calificaciones
- Estudio de Caso Sistema EndocrinoDocumento3 páginasEstudio de Caso Sistema EndocrinoAngelica Moreno100% (1)
- Micro Resumen Ciclo Ovarico y EndometrialDocumento4 páginasMicro Resumen Ciclo Ovarico y EndometrialCriisthii HernandezAún no hay calificaciones
- Ovarios y TesticuosDocumento58 páginasOvarios y TesticuosSolanny RosadoAún no hay calificaciones
- Wuolah Free Embrio Notas JuntasDocumento256 páginasWuolah Free Embrio Notas JuntasRafael DavidAún no hay calificaciones