También podría gustarte
- Resistencia Del Organismo A La Infección Ii: Inmunidad Específica E InespecíficaDocumento29 páginasResistencia Del Organismo A La Infección Ii: Inmunidad Específica E InespecíficaPedidos YaAún no hay calificaciones
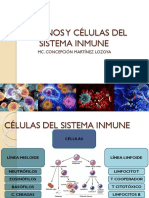
- Celulas Del Sistema InmuneDocumento33 páginasCelulas Del Sistema InmuneAngieAún no hay calificaciones
- Resumen Parcial de Biología PDFDocumento41 páginasResumen Parcial de Biología PDFcasidocmedicinaAún no hay calificaciones
- Datitos 1er Parcial PARTE 3Documento4 páginasDatitos 1er Parcial PARTE 3Agustina Palacios MachadoAún no hay calificaciones
- Celulas HematologicasDocumento38 páginasCelulas HematologicasGENETRIX451Aún no hay calificaciones
- Órganos Del Sistema LinfáticoDocumento44 páginasÓrganos Del Sistema LinfáticoNathalySinchiAún no hay calificaciones
- Resumen LeucocitosDocumento3 páginasResumen LeucocitosKelly VelaAún no hay calificaciones
- Equipo 4 "Linfopoyesis" ENF02A 20-2Documento16 páginasEquipo 4 "Linfopoyesis" ENF02A 20-2Ruth Cobo RosalesAún no hay calificaciones
- SISTEMA LINFOIDE-comprimidoDocumento55 páginasSISTEMA LINFOIDE-comprimido202220480Aún no hay calificaciones
- Inflamación Aguda - Equipo 2Documento51 páginasInflamación Aguda - Equipo 2Verónica Alejandra Pérez MoralesAún no hay calificaciones
- NeutrofilosDocumento6 páginasNeutrofilosMeleMeleninAún no hay calificaciones
- 04 Complemento, HLA, Presentacion AntigénicaDocumento46 páginas04 Complemento, HLA, Presentacion AntigénicaSilvana Mercedes Quintana CasusolAún no hay calificaciones
- Citocinas 2017Documento47 páginasCitocinas 2017Junior ValdiviezoAún no hay calificaciones
- URP Seminario 01 Filogenia Del Sistema InmuneDocumento25 páginasURP Seminario 01 Filogenia Del Sistema InmuneYébeny Tufino AlvaradoAún no hay calificaciones
- Tejido y Organos Lin 657041 Downloadable 1578071Documento10 páginasTejido y Organos Lin 657041 Downloadable 1578071calivilli5Aún no hay calificaciones
- Clase 2 Inmunidad Innata Parte IIDocumento9 páginasClase 2 Inmunidad Innata Parte IIfialophora0210Aún no hay calificaciones
- Órganos Linfáticos - InmunidadDocumento8 páginasÓrganos Linfáticos - InmunidadDanilo CavassimAún no hay calificaciones
- Semana 2 Circulación Leuco e Inmunidad InnataDocumento45 páginasSemana 2 Circulación Leuco e Inmunidad Innatam.ortega.quidelAún no hay calificaciones
- Parte3 Células Del Sistema InmuneDocumento31 páginasParte3 Células Del Sistema InmuneJose QuinteroAún no hay calificaciones
- PDF-t5 Respuesta InmunitariaDocumento32 páginasPDF-t5 Respuesta InmunitariamiriamAún no hay calificaciones
- INMUNOLOGIADocumento13 páginasINMUNOLOGIADiego SuntasigAún no hay calificaciones
- Linfoide 2Documento69 páginasLinfoide 2tomas563Aún no hay calificaciones
- PLAQUETASDocumento74 páginasPLAQUETASapi-3711117100% (2)
- Intercambio CelularDocumento4 páginasIntercambio CelularSalomé EstévezAún no hay calificaciones
- FagocitocisDocumento4 páginasFagocitocisRai GuerreroAún no hay calificaciones
- Clase 4 Los Neutrófilos y Sus ProductosDocumento35 páginasClase 4 Los Neutrófilos y Sus Productosvictor quispe Castro100% (1)
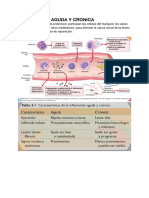
- Inflam Aguda y CronicaDocumento21 páginasInflam Aguda y CronicaMonica Sabater FrasquetAún no hay calificaciones
- Tema 18 Inmuno Dani RodríguezDocumento8 páginasTema 18 Inmuno Dani RodríguezDani RodriguezAún no hay calificaciones
- Órganos y Células Del Sistema InmuneDocumento50 páginasÓrganos y Células Del Sistema InmuneAngie ManningsAún no hay calificaciones
- Tema 0. PRODUCCIÓN, ESTRUCTURA Y FUNCIÓN DE LAS PLAQUETAS 2Documento17 páginasTema 0. PRODUCCIÓN, ESTRUCTURA Y FUNCIÓN DE LAS PLAQUETAS 2yuly zambranoAún no hay calificaciones
- Quimiocinas y Endotelio Vascular - MicDocumento29 páginasQuimiocinas y Endotelio Vascular - MicMiguel Ibañez CardenasAún no hay calificaciones
- Resumen Formas InespecificasDocumento8 páginasResumen Formas InespecificasSofAún no hay calificaciones
- MediadoresDocumento15 páginasMediadoresAndres Le FontiAún no hay calificaciones
- Inmunidad Adaptativa y Organos LinfoidesDocumento101 páginasInmunidad Adaptativa y Organos LinfoidesNAOMI BETZEL CASTRO ANDRADEAún no hay calificaciones
- Sesión 1 CelulasDocumento5 páginasSesión 1 CelulasDaniel Duran MoenaAún no hay calificaciones
- Seminario InmunoDocumento8 páginasSeminario InmunoMar Vidal LópezAún no hay calificaciones
- T Lymphocye Transmigration (Diapedesis de Linfocitos T)Documento1 páginaT Lymphocye Transmigration (Diapedesis de Linfocitos T)JostinWZAún no hay calificaciones
- 3 BasesanatomicasycelularesDocumento50 páginas3 BasesanatomicasycelularesElizabeth Román RamirezAún no hay calificaciones
- InflamaciDocumento74 páginasInflamaciYirianis LozadaAún no hay calificaciones
- Tejidos LinfoidesDocumento33 páginasTejidos LinfoidesMishel Jordan ZeballosAún no hay calificaciones
- t.4. - Sistema Linfoide - Dra. DeysiDocumento28 páginast.4. - Sistema Linfoide - Dra. DeysiEdgar Manuel Romero DíazAún no hay calificaciones
- Interacciones Entre El Leucocito y La Célula EndotelialDocumento4 páginasInteracciones Entre El Leucocito y La Célula EndotelialJamil Litardo VillacresesAún no hay calificaciones
- Medula OseaDocumento24 páginasMedula OseaMinerva García G.0% (1)
- Inflamacion Granulomatosa...Documento14 páginasInflamacion Granulomatosa...Thiago Mendes da SilvaAún no hay calificaciones
- Linfopoyesis IntroductorioDocumento37 páginasLinfopoyesis IntroductorioAna Claudia Bullon CuadraAún no hay calificaciones
- Clase 4-23 TMDocumento73 páginasClase 4-23 TMYuLi WAún no hay calificaciones
- Tema 5Documento18 páginasTema 5yelenabatista2Aún no hay calificaciones
- Cap. 81 TransplantesDocumento18 páginasCap. 81 TransplantesAbraham JimenezAún no hay calificaciones
- Tema 10 - Moleculas de Adhesión y Sus LigandosDocumento14 páginasTema 10 - Moleculas de Adhesión y Sus LigandosJaimeAún no hay calificaciones
- Tejido LinfoideDocumento5 páginasTejido LinfoideSeba Vallejos100% (1)
- Examen HistologiaDocumento8 páginasExamen HistologiaMario Orlando Chumioque SanchezAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Lentivirus 1Documento35 páginasLentivirus 1IVETH GABRIELA MENDEZ MURCIAAún no hay calificaciones
- ULTIMO Examen FisioDocumento8 páginasULTIMO Examen FisioIVETH GABRIELA MENDEZ MURCIAAún no hay calificaciones
- MOPECE3Documento96 páginasMOPECE3Alberto Rebaza71% (7)
- MOPECE1Documento34 páginasMOPECE1Andrea Luladasilva0% (1)
- Clamidias.Documento12 páginasClamidias.Heibyng MoranAún no hay calificaciones
- 14 - BrucelosisDocumento8 páginas14 - BrucelosisAbdeel MorazanAún no hay calificaciones
- Hemato Primera SemanaDocumento5 páginasHemato Primera SemanaBlanca HernándezAún no hay calificaciones
- LHPCTCCTomo IIIDocumento650 páginasLHPCTCCTomo IIIMiguelAngelBrocAún no hay calificaciones
- Antibiòticos de La Madre NaturalezaDocumento6 páginasAntibiòticos de La Madre Naturalezajorgecf123Aún no hay calificaciones
- Proyecto Steam Primera Semana 1Documento4 páginasProyecto Steam Primera Semana 1Domi TipazAún no hay calificaciones
- Colitis Ulcerosa y Enfermedad de CrohnDocumento19 páginasColitis Ulcerosa y Enfermedad de CrohnErnesto Enrique Garcia VasquezAún no hay calificaciones
- S3 MC Endotelio VascularDocumento1 páginaS3 MC Endotelio VascularRoberth Dianiro Sanchez RodriguezAún no hay calificaciones
- INMUNONUTRICIONDocumento6 páginasINMUNONUTRICIONmatacurieAún no hay calificaciones
- Clase 5 Inmunoprofilaxis e Inmunoterapia-Reacciones de HipersensibilidadDocumento47 páginasClase 5 Inmunoprofilaxis e Inmunoterapia-Reacciones de HipersensibilidadJose Luis Tomateo ValenciaAún no hay calificaciones
- Caso Clinico - Ácaros - ExpoDocumento25 páginasCaso Clinico - Ácaros - ExpoMilly RivasAún no hay calificaciones
- 5 Tipos de Glóbulos Blancos en La Sangre PeriféricaDocumento5 páginas5 Tipos de Glóbulos Blancos en La Sangre PeriféricaUriel BriceñoAún no hay calificaciones
- Paramixovirus y Otros Virus Respiratorios - 2016Documento32 páginasParamixovirus y Otros Virus Respiratorios - 2016Jerónimo Marcía CervantesAún no hay calificaciones
- Tipos de Leche Por El Sistema de HigienizaciónDocumento7 páginasTipos de Leche Por El Sistema de HigienizaciónCristhian Tasayco100% (1)
- Leche de CabraDocumento32 páginasLeche de CabraDino Martin Velasquez GalindoAún no hay calificaciones
- Inform3 de Biologia Celulas Sanguineas.Documento6 páginasInform3 de Biologia Celulas Sanguineas.Lina LarrotaAún no hay calificaciones
- Examen Grado 9 4 PeriodoDocumento6 páginasExamen Grado 9 4 Periodorosaaide villalbaAún no hay calificaciones
- Manejo Corioamnionitis v.1Documento12 páginasManejo Corioamnionitis v.1Saudith Charris EscorciaAún no hay calificaciones
- Laboratorio de Analisis Clinicos: Av. Instituto Politécnico Nacional 5160 Magdalena de Las Salinas TEL 5577920632Documento4 páginasLaboratorio de Analisis Clinicos: Av. Instituto Politécnico Nacional 5160 Magdalena de Las Salinas TEL 5577920632Suilevrom MorveliusAún no hay calificaciones
- Alergias AlimentariasDocumento3 páginasAlergias AlimentariasMagali CoroAún no hay calificaciones
- MFPH II CuestionarioDocumento34 páginasMFPH II CuestionarioRubén Rivas100% (1)
- Toxicología en LechugasDocumento9 páginasToxicología en LechugasMauricio VidalAún no hay calificaciones
- Mapa AplvDocumento1 páginaMapa AplvMvm LakmirAún no hay calificaciones
- Silabo de Patología Clínica y Transfusional 2018-Ii - 20180806114337Documento17 páginasSilabo de Patología Clínica y Transfusional 2018-Ii - 20180806114337Johanna Lissethe Velasquez HerrerosAún no hay calificaciones
- HematopoyesisDocumento1 páginaHematopoyesisRoderick Miranda100% (1)
- Cetirizine (Systemic) - Drug Information - UpToDateDocumento33 páginasCetirizine (Systemic) - Drug Information - UpToDateJosué Pablo Chicaiza AbadAún no hay calificaciones
- CAPÍTULO 12 - Respuestas Efectoras - Inmunidad Mediada Por Anticuerpos y Por CélulasDocumento54 páginasCAPÍTULO 12 - Respuestas Efectoras - Inmunidad Mediada Por Anticuerpos y Por CélulasPablo Osvaldo Gómez SilvaAún no hay calificaciones
- Manifestaciones Clinicas Del SidaDocumento4 páginasManifestaciones Clinicas Del SidaEnrique MartinezAún no hay calificaciones
- Anticuerpos Monoclonales-Articulo1Documento78 páginasAnticuerpos Monoclonales-Articulo1Reichell SanchezAún no hay calificaciones
- Evaluaciones 3ra. UnidadDocumento9 páginasEvaluaciones 3ra. UnidadYan RavanalesAún no hay calificaciones
- Catálogo Ecuador 4lifeDocumento29 páginasCatálogo Ecuador 4lifeSEN MARCOS PEREIRAAún no hay calificaciones
- F4e1f15325f0d7d1 Fusionado PDFDocumento454 páginasF4e1f15325f0d7d1 Fusionado PDFEnrique Quispe Quito100% (1)