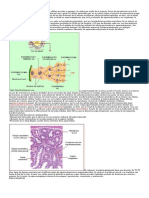
También podría gustarte
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Equinotecnia - 2011Documento26 páginasEquinotecnia - 2011Repositorio de la Biblioteca de la DEA-MAG-PYAún no hay calificaciones
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Ovogenesis y EspermatogénesisDocumento9 páginasOvogenesis y EspermatogénesisManuel Rosado100% (1)
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Sistema Endocrino para Quinto de Primaria PDFDocumento5 páginasSistema Endocrino para Quinto de Primaria PDFJill Halinna Terán Montalván100% (4)
- Calvo y CabreadoDocumento49 páginasCalvo y CabreadoOscar EstoaAún no hay calificaciones
- EmbriologíaDocumento14 páginasEmbriologíaIldefonso Mendoza Rueda100% (1)
- Prueba Ciencias Naturales 6to Básicos Sistema ReproductorDocumento6 páginasPrueba Ciencias Naturales 6to Básicos Sistema ReproductorDaniela Pineda100% (1)
- Conocimientos y actitudes sexuales adolescentesDocumento94 páginasConocimientos y actitudes sexuales adolescentesJc Ca0% (2)
- Aparato Reproductor Del MachoDocumento10 páginasAparato Reproductor Del MachoVanne Ag100% (1)
- UABP 2, 6 Y 9 - EmbriologíaDocumento15 páginasUABP 2, 6 Y 9 - EmbriologíaGiovanni MedranoAún no hay calificaciones
- Qué Es La GametogénesisDocumento3 páginasQué Es La GametogénesisRubenAún no hay calificaciones
- Gametognesis BiologiaDocumento15 páginasGametognesis BiologiaJosue RebolledoAún no hay calificaciones
- Guía 2do Parcial de Histología, Embriología y GenéticaDocumento17 páginasGuía 2do Parcial de Histología, Embriología y GenéticaKim ZoldyckAún no hay calificaciones
- EspermatogénesisDocumento5 páginasEspermatogénesisXavier GuillenAún no hay calificaciones
- Marco Teórico: Generalidades Del Aparato Reproductor MasculinoDocumento18 páginasMarco Teórico: Generalidades Del Aparato Reproductor MasculinoAndrea IriasAún no hay calificaciones
- El Sistema Genital MasculinoDocumento11 páginasEl Sistema Genital MasculinoKarla Priscila AndradeAún no hay calificaciones
- EspermatogenesisDocumento5 páginasEspermatogenesisDiana Jimenez MartinezAún no hay calificaciones
- TesticuloDocumento35 páginasTesticuloLaura Angelica de PerezAún no hay calificaciones
- Fundamentos Dela Reproducción Humana.Documento10 páginasFundamentos Dela Reproducción Humana.Iris MendozaAún no hay calificaciones
- Embriologia Falta CaratulaDocumento40 páginasEmbriologia Falta CaratulaSaira Diaz AldanaAún no hay calificaciones
- GAMETOGÉNESISDocumento7 páginasGAMETOGÉNESISMiguel Angel Parra GarcíaAún no hay calificaciones
- OvogénesisDocumento5 páginasOvogénesisKevin CisAún no hay calificaciones
- Resumen de Embriologia LangmanDocumento4 páginasResumen de Embriologia LangmanMelanie GrantonAún no hay calificaciones
- Fisiología de GametogénesisDocumento5 páginasFisiología de Gametogénesispier manuel cabezas armellonAún no hay calificaciones
- Marco TeóricoDocumento2 páginasMarco TeóricoKatherine A. Mendoza De La CruzAún no hay calificaciones
- Primera Semana Del Desarrollo Humano Capítulo 2Documento12 páginasPrimera Semana Del Desarrollo Humano Capítulo 2isaAún no hay calificaciones
- EMI MonografiaDocumento25 páginasEMI MonografiaAlejandro Mendoza SarmientoAún no hay calificaciones
- EmbriologíaDocumento17 páginasEmbriologíaCarmen Porras Jarufe50% (2)
- Espermatogénesis y OvogenesisDocumento25 páginasEspermatogénesis y OvogenesisIsrael Silva SaucedoAún no hay calificaciones
- GAMETOGENÉSISDocumento6 páginasGAMETOGENÉSISanon_757572911Aún no hay calificaciones
- Espermatogénesis: Formación de EspermatozoidesDocumento9 páginasEspermatogénesis: Formación de EspermatozoidesJasminii Flor Llatas MontenegroAún no hay calificaciones
- EMBRIOLOGIADocumento9 páginasEMBRIOLOGIAAlejandra MartinezAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento11 páginasAparato Reproductor MasculinoNair MartínezAún no hay calificaciones
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 páginasEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezAún no hay calificaciones
- Tema 1 Desarrollo EmbrionarioDocumento19 páginasTema 1 Desarrollo EmbrionarioAnder GroundAún no hay calificaciones
- Introduccion a la EmbriologiaDocumento24 páginasIntroduccion a la EmbriologiaMARGARET MICHELLE CABREJA ROSARIOAún no hay calificaciones
- Resumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital MasculinoDocumento13 páginasResumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital Masculinovictoriaetchegoyen123Aún no hay calificaciones
- ESPERMATOGENESISDocumento15 páginasESPERMATOGENESISMiriam E. MairimAún no hay calificaciones
- E SpermatogenesisDocumento6 páginasE SpermatogenesisThalía YanchapaxiAún no hay calificaciones
- EmbrioDocumento231 páginasEmbrioOmar C. RodríguezAún no hay calificaciones
- Gametogénesis y ciclo sexual.docxDocumento4 páginasGametogénesis y ciclo sexual.docxValentina BOLDRINIAún no hay calificaciones
- Proceso GametogénesisDocumento4 páginasProceso GametogénesisAlexander EztradaAún no hay calificaciones
- Sistema ResumidoDocumento24 páginasSistema ResumidoJorge RamirezAún no hay calificaciones
- GametogenesisDocumento10 páginasGametogenesisSigmaSAún no hay calificaciones
- EmbriologíaDocumento24 páginasEmbriologíaDaniAún no hay calificaciones
- Apuntes embriologíaDocumento9 páginasApuntes embriologíasebastianAún no hay calificaciones
- GametogénesisDocumento7 páginasGametogénesisMonics GonzalezAún no hay calificaciones
- Fecundación, Desarrollo Embrionario y FetalDocumento11 páginasFecundación, Desarrollo Embrionario y FetalAmélie CastañoAún no hay calificaciones
- Espermatogenesis - TrabajoDocumento12 páginasEspermatogenesis - TrabajoJanella Tineo MedinaAún no hay calificaciones
- GAMETOGÉNESISDocumento3 páginasGAMETOGÉNESISMartina GentilAún no hay calificaciones
- EspermatogénesisDocumento3 páginasEspermatogénesisSteveAún no hay calificaciones
- GAMETOGENESIS ExposicionDocumento5 páginasGAMETOGENESIS ExposicionKEITLYN BENTHAMS SILVA100% (1)
- Formación de gametos masculinos y femeninos (espermatozoides y óvulosDocumento9 páginasFormación de gametos masculinos y femeninos (espermatozoides y óvulosAixa CamposAún no hay calificaciones
- Selem Channan Trabajo de FisiologíaDocumento7 páginasSelem Channan Trabajo de FisiologíaKenary GonzalezAún no hay calificaciones
- Resumen Final Embriología.Documento17 páginasResumen Final Embriología.Nazareth PacayutAún no hay calificaciones
- Gametogénesis: Formación de GametosDocumento28 páginasGametogénesis: Formación de GametosSofia PantojaAún no hay calificaciones
- 17. Aparato Reproductor Masculino 1Documento8 páginas17. Aparato Reproductor Masculino 1malubonfigliAún no hay calificaciones
- Sistema Reproductor MasculinoDocumento135 páginasSistema Reproductor MasculinoPaola Azucena SalasAún no hay calificaciones
- Mitosis Meiosis 1Documento37 páginasMitosis Meiosis 1Alexander b.Aún no hay calificaciones
- 3 Embriología Del Aparato Genital FemeninoDocumento24 páginas3 Embriología Del Aparato Genital FemeninoluceroAún no hay calificaciones
- En Qué Consiste La GametogenesisDocumento3 páginasEn Qué Consiste La GametogenesisDarioAún no hay calificaciones
- SISTEMA ENDOCRINO TareaDocumento1 páginaSISTEMA ENDOCRINO TareaCAMILA MARTINEZ GALLEGOS 2 SECUNDARIAAún no hay calificaciones
- Organo Genital Masculino y FemeninoDocumento38 páginasOrgano Genital Masculino y FemeninoRomell Morales VilchezAún no hay calificaciones
- 1ra Evaluación Trimestral. 5° (24 - OCT AL 1° NOV.)Documento25 páginas1ra Evaluación Trimestral. 5° (24 - OCT AL 1° NOV.)Pedro CamejoAún no hay calificaciones
- 06 STR #06 Final Usamedic 2017 Print Alu PDFDocumento24 páginas06 STR #06 Final Usamedic 2017 Print Alu PDFkira tomokiAún no hay calificaciones
- Embriologia CompletooDocumento27 páginasEmbriologia CompletooMagui Esquivel100% (1)
- Prueba de Comprensión de La Naturaleza "Sistema Reproductor Masculino"Documento2 páginasPrueba de Comprensión de La Naturaleza "Sistema Reproductor Masculino"IgnacioRojas100% (1)
- Exposicion GinecoDocumento39 páginasExposicion GinecoLizbet Jasmin RodríguezAún no hay calificaciones
- Varicoccele (Autoguardado)Documento28 páginasVaricoccele (Autoguardado)ZENTENOAún no hay calificaciones
- Silabo Anatomia y Fisiologia IIDocumento11 páginasSilabo Anatomia y Fisiologia IIARNOLD ALFAROAún no hay calificaciones
- Evacuación de Gametos de Lapa Frutilla - Fissurella Cumingi Reeve, 1849 - y Lapa NegraDocumento64 páginasEvacuación de Gametos de Lapa Frutilla - Fissurella Cumingi Reeve, 1849 - y Lapa NegraLuis Gallardo Barraza100% (1)
- ESPERMATOGENESISDocumento39 páginasESPERMATOGENESISJUAN JOSEAún no hay calificaciones
- Prototipo Electroeyaculador para Aves en Peligro de Extinción en El SalvadorDocumento5 páginasPrototipo Electroeyaculador para Aves en Peligro de Extinción en El SalvadorLuis BarriereAún no hay calificaciones
- Apuntes 1Documento20 páginasApuntes 1mariaamachucamAún no hay calificaciones
- Crecimiento y desarrollo del escolar y adolescenteDocumento34 páginasCrecimiento y desarrollo del escolar y adolescenteBruno Daniel OliveraAún no hay calificaciones
- Atlas de Imagenes Patologicas CorregidoDocumento44 páginasAtlas de Imagenes Patologicas CorregidoMarcelo MartínezAún no hay calificaciones
- Criptorquidia y ectopia testicular: causas, diagnóstico y tratamientoDocumento6 páginasCriptorquidia y ectopia testicular: causas, diagnóstico y tratamientoMaritza GómezAún no hay calificaciones
- VaricoceleDocumento2 páginasVaricoceleYALIELAún no hay calificaciones
- Cáncer Testicular: Factores, Tipos y TratamientoDocumento23 páginasCáncer Testicular: Factores, Tipos y TratamientoMlm Glez100% (1)
- 002-Fisiología y Bioquímica de La Hembra y El MachoDocumento48 páginas002-Fisiología y Bioquímica de La Hembra y El MachoJazmin VeraAún no hay calificaciones
- MarielysDocumento28 páginasMarielysAdri LopezAún no hay calificaciones
- Criptorquidia - JGMGDocumento1 páginaCriptorquidia - JGMGJ Gerardo GarciaAún no hay calificaciones
- Genitales MasculinosDocumento71 páginasGenitales MasculinosProfesor Daniel Da RosaAún no hay calificaciones
- Reproduccion AnimalDocumento32 páginasReproduccion AnimalMaría Gabriela ArangurenAún no hay calificaciones
- Funciones Reproductoras y Hormonales MasculinasDocumento25 páginasFunciones Reproductoras y Hormonales MasculinasPedro Miguel EgüezAún no hay calificaciones