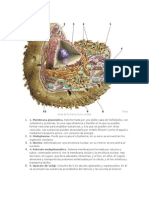
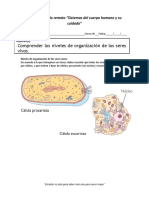
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Separata Organelas y Componentes Extracelulares de Una Celula EucariotaDocumento9 páginasSeparata Organelas y Componentes Extracelulares de Una Celula Eucariotalucero liraAún no hay calificaciones
- Biologia Celular TrabajoDocumento30 páginasBiologia Celular TrabajoSaraiOlmedoCruzAún no hay calificaciones
- La célula: estructura y funciones de sus orgánulosDocumento9 páginasLa célula: estructura y funciones de sus orgánulosDavid MosqueraAún no hay calificaciones
- La CelulaDocumento4 páginasLa CelulaKarolyne RealAún no hay calificaciones
- Gartner, Leslie P. - Texto Atlas de Histologia, 2da Edición (2 Citoplasma)Documento35 páginasGartner, Leslie P. - Texto Atlas de Histologia, 2da Edición (2 Citoplasma)Rocío Ruiz100% (4)
- Apuntes Biologia CelularDocumento31 páginasApuntes Biologia CelularNoelia FernándzAún no hay calificaciones
- Célula Vegetal 2013 PDFDocumento26 páginasCélula Vegetal 2013 PDFivan villarrealAún no hay calificaciones
- Tema 6 - El Citoplasma CelularDocumento4 páginasTema 6 - El Citoplasma CelularEmma MiñanaAún no hay calificaciones
- Estructura Biologica y Funcional Del Ser Humano PDFDocumento14 páginasEstructura Biologica y Funcional Del Ser Humano PDFJulioCarlosMartinAún no hay calificaciones
- Guía Célula y FuncionesDocumento2 páginasGuía Célula y FuncionesJasna ZovakAún no hay calificaciones
- 3 Cuadernillo. Características de Las CélulasDocumento7 páginas3 Cuadernillo. Características de Las Célulasa353862Aún no hay calificaciones
- Células Eucariotas y ProcariotasDocumento5 páginasCélulas Eucariotas y ProcariotasFanny CMunvAún no hay calificaciones
- Ensayo Lc-034 2017 - ProDocumento16 páginasEnsayo Lc-034 2017 - ProBrianda RuizAún no hay calificaciones
- Apunte de Biologia Fundamentos de La BioDocumento31 páginasApunte de Biologia Fundamentos de La BioRon RuizAún no hay calificaciones
- Cuestionario Estructura Celular, Membranas y TransporteDocumento7 páginasCuestionario Estructura Celular, Membranas y TransporteloinconcienteAún no hay calificaciones
- Celula Animal BioDocumento6 páginasCelula Animal BioAmor MiraveteAún no hay calificaciones
- Unidad I-1-1Documento31 páginasUnidad I-1-1Katerin VenturaAún no hay calificaciones
- Resumen Biología Examen 18 - 01Documento13 páginasResumen Biología Examen 18 - 01adnAún no hay calificaciones
- Clase 5Documento75 páginasClase 5Yesenia PabonAún no hay calificaciones
- Celula AnimaDocumento5 páginasCelula Animapo poAún no hay calificaciones
- CÉLULADocumento9 páginasCÉLULARebeca lanauAún no hay calificaciones
- Citoplasma CelularDocumento19 páginasCitoplasma CelularAlisson Caicedo100% (2)
- Fisiología TP1 PDFDocumento5 páginasFisiología TP1 PDFMilena OteroAún no hay calificaciones
- Célula AnimalDocumento7 páginasCélula AnimalRolex Gonzalo100% (1)
- Tema 1. Organización General Del Cuerpo Humano. AnatomíaDocumento13 páginasTema 1. Organización General Del Cuerpo Humano. AnatomíalolisocialmediaAún no hay calificaciones
- Guía de Organelos CelularesDocumento8 páginasGuía de Organelos CelularesCamila Bossio CastillaAún no hay calificaciones
- La célula: estructura y funciones básicasDocumento6 páginasLa célula: estructura y funciones básicasYusiley Zeballos MejiaAún no hay calificaciones
- Actividad de Repaso Procesos Biologicos OctavoDocumento7 páginasActividad de Repaso Procesos Biologicos OctavoSANDRAAún no hay calificaciones
- Sistema de EndomembranasDocumento9 páginasSistema de Endomembranasandrea rochaAún no hay calificaciones
- La CélulaDocumento26 páginasLa CélulaGabrielaAún no hay calificaciones
- Organelas celulares: funciones y presenciaDocumento9 páginasOrganelas celulares: funciones y presenciaMicaelaAún no hay calificaciones
- Sistema de EndomembranasDocumento53 páginasSistema de EndomembranasMarcelo SuarezAún no hay calificaciones
- Unidad IDocumento31 páginasUnidad IErick CuauroAún no hay calificaciones
- CELULA EUCARIOTADocumento25 páginasCELULA EUCARIOTAvalentinaquispe102Aún no hay calificaciones
- Taller 5 Membrana-Citoesqueleto EndomembranasDocumento5 páginasTaller 5 Membrana-Citoesqueleto EndomembranasDaniela CifuentesAún no hay calificaciones
- Resumo CÉLULASDocumento3 páginasResumo CÉLULASEllen SchaeferAún no hay calificaciones
- Citoplasma y Organelas - Segundo Año.Documento6 páginasCitoplasma y Organelas - Segundo Año.estebanrodriguez9084uAún no hay calificaciones
- La CélulaDocumento42 páginasLa CélulaEduardo Asael Tlaxalo RamirezAún no hay calificaciones
- Fundamentos de La Célula y El CitoplasmaDocumento5 páginasFundamentos de La Célula y El CitoplasmaLuis InfanteAún no hay calificaciones
- Tema 3 Estructura y Función CelularDocumento8 páginasTema 3 Estructura y Función CelularJorge GarcíaAún no hay calificaciones
- U2 Act 2 Estructura Fisica de La CelulaDocumento15 páginasU2 Act 2 Estructura Fisica de La CelulaKAROL GUTIERREZ MADRIGALAún no hay calificaciones
- Final PsicofisiologiaDocumento2 páginasFinal PsicofisiologiaMicaela AguileraAún no hay calificaciones
- Ultraestructura de MembranaDocumento4 páginasUltraestructura de MembranaDIANA CAROLINA PEREZ ACOSTAAún no hay calificaciones
- Ud 2 La Célula Apuntes ExamenDocumento17 páginasUd 2 La Célula Apuntes ExamenMaria EgeaAún no hay calificaciones
- La Celula y Elementos ConstitutivosDocumento45 páginasLa Celula y Elementos ConstitutivosBeth DiazAún no hay calificaciones
- BIO - Sem4 - Citología IIDocumento10 páginasBIO - Sem4 - Citología IIJadira LlerenaAún no hay calificaciones
- Conferencia Citoplasma Completo OkDocumento60 páginasConferencia Citoplasma Completo OkPawan Alvarez PerezAún no hay calificaciones
- Célula: estructura y tiposDocumento30 páginasCélula: estructura y tiposnanciAún no hay calificaciones
- TEMACÉLULADocumento4 páginasTEMACÉLULAHUGO CESAR QUIROZ SUAREZAún no hay calificaciones
- Redaccion de Expo de BioDocumento6 páginasRedaccion de Expo de BioGabriela RamírezAún no hay calificaciones
- MÓDULO 4 (Células) - BIOLOGIA (Curso de Ingreso UnLaM)Documento7 páginasMÓDULO 4 (Células) - BIOLOGIA (Curso de Ingreso UnLaM)Luisa GuerreroAún no hay calificaciones
- Organelos CelularesDocumento38 páginasOrganelos Celularesyoselin gonzalezAún no hay calificaciones
- Partes de La CelulaDocumento1 páginaPartes de La CelulaOsmandy VeraAún no hay calificaciones
- Botánica, CitologíaDocumento10 páginasBotánica, CitologíaBárbara SmerkinAún no hay calificaciones
- Resumenes de Histología: Citología y Tejido EpitelialDocumento18 páginasResumenes de Histología: Citología y Tejido EpitelialErica Monterrey MoraAún no hay calificaciones
- Guia 1 Anatomia CelularDocumento10 páginasGuia 1 Anatomia CelularGloriaAún no hay calificaciones
- El Esquema de La Estructura de Una Célula AnimalDocumento4 páginasEl Esquema de La Estructura de Una Célula AnimalCyndi Martinez D' Codeshka0% (1)
- Bases Biolgicas ResumenDocumento2 páginasBases Biolgicas ResumenValentina Torres SAún no hay calificaciones
- 4 Alimentaciónnaturalezaycultura PDFDocumento10 páginas4 Alimentaciónnaturalezaycultura PDFNataliaAún no hay calificaciones
- Tema 5 - Compuestos de Carbono - BQE2021Documento23 páginasTema 5 - Compuestos de Carbono - BQE2021NataliaAún no hay calificaciones
- Importancia biológica del agua: Estructura molecular, propiedades y funcionesDocumento24 páginasImportancia biológica del agua: Estructura molecular, propiedades y funcionesNataliaAún no hay calificaciones
- Práctica 2 Electroforesis - 2021Documento3 páginasPráctica 2 Electroforesis - 2021NataliaAún no hay calificaciones
- P1 - 2021 - Introducción Laboratorio - Curva de Valoración de Un Aminoácido PDFDocumento6 páginasP1 - 2021 - Introducción Laboratorio - Curva de Valoración de Un Aminoácido PDFNataliaAún no hay calificaciones
- Clase 5 BioenergéticaDocumento78 páginasClase 5 BioenergéticaMANUEL ANDRES VILLANUEVA LEONAún no hay calificaciones
- UDLA - BiomoléculasDocumento25 páginasUDLA - BiomoléculasKIMBERLY ANNAYS SANDOVAL ROMEROAún no hay calificaciones
- 1esocnc2 Ev Esu1Documento2 páginas1esocnc2 Ev Esu1Isabel Maria Aguilar MorenoAún no hay calificaciones
- El Comportamiento AnimalDocumento5 páginasEl Comportamiento AnimalManuel OrihuelaAún no hay calificaciones
- Estudio de Macroinvertebrados en Los Ríos Alambí y PichanDocumento14 páginasEstudio de Macroinvertebrados en Los Ríos Alambí y PichanJoss TonatoAún no hay calificaciones
- Respuesta Taller #1 CienciasDocumento3 páginasRespuesta Taller #1 Cienciasmaget 7890Aún no hay calificaciones
- Tema 1: Captación Y Transformación de La Energía de La CélulaDocumento9 páginasTema 1: Captación Y Transformación de La Energía de La CélulaOlmedo QuinteroAún no hay calificaciones
- Crucigrama de BiodiversidadDocumento1 páginaCrucigrama de BiodiversidadSandra Saldaña100% (4)
- 2 Bach ByGDocumento34 páginas2 Bach ByGÁngela Díaz GarcíaAún no hay calificaciones
- Identifica biomoléculas en alimentosDocumento5 páginasIdentifica biomoléculas en alimentosCristopher García AyalaAún no hay calificaciones
- Agua, Hombre y PaisajeDocumento261 páginasAgua, Hombre y PaisajeCecilia Briones100% (1)
- Las PlantasDocumento33 páginasLas PlantasTFPAún no hay calificaciones
- Unidad 1 NeuroDocumento5 páginasUnidad 1 NeuroJavieer Aleejandro Estay FigueroaAún no hay calificaciones
- Genetica y AmbienteDocumento7 páginasGenetica y AmbienteNatalia Serrano CardozoAún no hay calificaciones
- Guía Curricular de BiologíaDocumento72 páginasGuía Curricular de BiologíajavierAún no hay calificaciones
- Normas Vitales y Normas Sociales - Cnaguilhem. PPT Andrea BenvenutoDocumento16 páginasNormas Vitales y Normas Sociales - Cnaguilhem. PPT Andrea BenvenutoVerónica ChiapelloAún no hay calificaciones
- Cuestionario de BiologíaDocumento5 páginasCuestionario de BiologíaCharlottAún no hay calificaciones
- Técnicas de Higienización, Manejo y Conservación deDocumento31 páginasTécnicas de Higienización, Manejo y Conservación deGusy HernandezAún no hay calificaciones
- Autoconciencia Por El MovDocumento23 páginasAutoconciencia Por El MovMaria AlejandraAún no hay calificaciones
- Los Cinco ReinosDocumento11 páginasLos Cinco Reinoshectordayan20% (1)
- Botanica II - Ensayo - Daniela Cabrera - Daniela - Cabrerag@ug - Edu.ecDocumento4 páginasBotanica II - Ensayo - Daniela Cabrera - Daniela - Cabrerag@ug - Edu.ecDaniela13519Aún no hay calificaciones
- Actividad 2 SexualDocumento2 páginasActividad 2 SexualOrlando RdlAún no hay calificaciones
- Prueba de Fotosíntesis 1 MedioDocumento5 páginasPrueba de Fotosíntesis 1 MedioLeila Beatriz Hidalgo Gamboa100% (1)
- Plan de Estudios Quimica Con CompetenciasDocumento28 páginasPlan de Estudios Quimica Con CompetenciasHéctor Wyllian Melo MartínezAún no hay calificaciones
- Guia 1 - Ciencias Pie OctavoDocumento6 páginasGuia 1 - Ciencias Pie OctavoManuel ParraAún no hay calificaciones
- Guíaradio SecundariaDocumento59 páginasGuíaradio Secundariadperalta30100% (1)
- Tarea II PsicologiaDocumento8 páginasTarea II PsicologiaIdeas CenterAún no hay calificaciones
- EQUIPO 4 - PÍA - BioSalDocumento6 páginasEQUIPO 4 - PÍA - BioSalJulisa HernandezAún no hay calificaciones
- Funciones de Relación en Los Seres VivosDocumento2 páginasFunciones de Relación en Los Seres VivosMarcko's SalazarAún no hay calificaciones
- Funciones VitalesDocumento2 páginasFunciones VitalesDavid FRANCO FARFÁNAún no hay calificaciones