También podría gustarte
- Anatomía y Fisiología CelularDocumento7 páginasAnatomía y Fisiología CelularDALVIC ALARCON50% (2)
- Reticulo Endoplasmatico Liso y Rugoso, FuncionesDocumento5 páginasReticulo Endoplasmatico Liso y Rugoso, FuncionesAna MarcelaAún no hay calificaciones
- Componentes de La CelulaDocumento5 páginasComponentes de La CelulaMarco Antonio Quiroga BustinzaAún no hay calificaciones
- CemorDocumento8 páginasCemorclaudioAún no hay calificaciones
- CELULADocumento108 páginasCELULAalejandraAún no hay calificaciones
- Marco TeóricoDocumento6 páginasMarco TeóricoEdison OrtizAún no hay calificaciones
- Citoplasma y Reticulo EndoplasmaticoDocumento13 páginasCitoplasma y Reticulo EndoplasmaticoNoemi2466Aún no hay calificaciones
- Biologia Tema 07Documento6 páginasBiologia Tema 07sylvanaAún no hay calificaciones
- Organelos CitoplasmáticosDocumento24 páginasOrganelos CitoplasmáticosAlbaniBriceñoAún no hay calificaciones
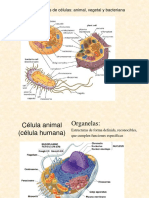
- Célula AnimalDocumento17 páginasCélula AnimalJUDITH ALONDRA TORRES HERNANDEZAún no hay calificaciones
- BiologiaDocumento156 páginasBiologiaKatherine AltamiranoAún no hay calificaciones
- RibosomasDocumento6 páginasRibosomaspakalna80% (5)
- Las CélulasDocumento5 páginasLas CélulasMarta Zapatera SansAún no hay calificaciones
- Cito PlasmaDocumento8 páginasCito PlasmaWalter oliveraAún no hay calificaciones
- Apunte La..Documento11 páginasApunte La..Guillermina ArroyoAún no hay calificaciones
- MitocondriasDocumento6 páginasMitocondriasemmanuelsojuelsilvaAún no hay calificaciones
- Resumen de Biologia Celular - Agosto.2021Documento8 páginasResumen de Biologia Celular - Agosto.2021Michell AndradeAún no hay calificaciones
- Celula Animal y VegetalDocumento7 páginasCelula Animal y VegetalClaudia MarquezAún no hay calificaciones
- Unidad 3 - La CélulaDocumento16 páginasUnidad 3 - La Célulaalexis menjibarAún no hay calificaciones
- Semianual Biologia Tema 4Documento9 páginasSemianual Biologia Tema 4Friedrich Wilhelm NietzscheAún no hay calificaciones
- Resumen Capítulo 2 Del Libro de HistologíaDocumento56 páginasResumen Capítulo 2 Del Libro de HistologíaMedicina BlogAún no hay calificaciones
- Organitos CitoplasmaticosDocumento4 páginasOrganitos CitoplasmaticosMartAún no hay calificaciones
- La Estructura de La CélulaDocumento6 páginasLa Estructura de La CélulaMaricielo Calderon MinayaAún no hay calificaciones
- Bioquimica Vet 1Documento15 páginasBioquimica Vet 1FELIX ALEX CONDORI NINAAún no hay calificaciones
- Celula VegetalDocumento31 páginasCelula Vegetaljheinercarias27Aún no hay calificaciones
- El Citoplasma BiologiaDocumento6 páginasEl Citoplasma BiologiaSteeven Espinosa OtavaloAún no hay calificaciones
- Guia de Apoyo BioquimicaDocumento8 páginasGuia de Apoyo BioquimicaCarla Cecilia Gonzalez SuberoAún no hay calificaciones
- Biología Unidad 5-Estructura y Función CelularDocumento11 páginasBiología Unidad 5-Estructura y Función CelularJenny MoreiraAún no hay calificaciones
- Membrana Plasmã¡tica EucariotaDocumento5 páginasMembrana Plasmã¡tica EucariotaJULISSA GONZÁLEZ VARGASAún no hay calificaciones
- Los Orgánulos Celulares Y Sus Funciones: Alba Garín Pes 1º BCS (Sástago)Documento9 páginasLos Orgánulos Celulares Y Sus Funciones: Alba Garín Pes 1º BCS (Sástago)Cesar Emmanuel Miss SalgadoAún no hay calificaciones
- Celulas InformeDocumento13 páginasCelulas InformeJose Banban88% (8)
- Organelas CelularesDocumento33 páginasOrganelas CelularesTamara ZabalaAún no hay calificaciones
- Continue: Organelos de La Célula Animal PDFDocumento3 páginasContinue: Organelos de La Célula Animal PDFJeanaime SamyAún no hay calificaciones
- Seg Exa BiologiaDocumento36 páginasSeg Exa BiologiaBrayan Villavicencio TovarAún no hay calificaciones
- 3-La CelulaDocumento5 páginas3-La CelulaVICTOR FUENTESAún no hay calificaciones
- BIOLOGÍA Tema 9 (Bloque 2)Documento7 páginasBIOLOGÍA Tema 9 (Bloque 2)Carolina De la peña fernandezAún no hay calificaciones
- Reticulo EndoplasmaticoDocumento5 páginasReticulo Endoplasmaticointer celAún no hay calificaciones
- Citoplasma de La Celula EucariotaDocumento12 páginasCitoplasma de La Celula Eucariotaleo apazaAún no hay calificaciones
- Semana 06 ESTRUCTURA Y FUNCION CELULARDocumento14 páginasSemana 06 ESTRUCTURA Y FUNCION CELULARMARKO ANTONIO RAMOS QUISPEAún no hay calificaciones
- Sistema de EndomembranasDocumento16 páginasSistema de EndomembranasAracelyIssaEnriquezTorrezAún no hay calificaciones
- Citoplasma CelularDocumento4 páginasCitoplasma CelularaxelAún no hay calificaciones
- Las Entidades Fundamentales de Todo Ser Vivo Son Las CélulasDocumento5 páginasLas Entidades Fundamentales de Todo Ser Vivo Son Las CélulasroximarcontrerasAún no hay calificaciones
- El Retículo EndoplasmáticoDocumento11 páginasEl Retículo EndoplasmáticoMartin CrisantoAún no hay calificaciones
- Resumen, Capítulo 2Documento12 páginasResumen, Capítulo 2MITZY YATZYRI VICTOR ARAUJOAún no hay calificaciones
- Fisiologia FinalDocumento7 páginasFisiologia FinalDaniela VelascoAún no hay calificaciones
- La Membrana CitoplasmaticaDocumento9 páginasLa Membrana CitoplasmaticaMilagros Gianella Cueva ArteagaAún no hay calificaciones
- Organelas Completo 2022Documento14 páginasOrganelas Completo 2022Sergio NarvaezAún no hay calificaciones
- Apuntes La Celula VegetalDocumento6 páginasApuntes La Celula VegetalKarenAún no hay calificaciones
- Eucariota, Organelas, Citoplasma, CitoesqueletoDocumento30 páginasEucariota, Organelas, Citoplasma, CitoesqueletoAlan GanemAún no hay calificaciones
- Membrana CelularDocumento6 páginasMembrana CelularKenlly Cutin CastilloAún no hay calificaciones
- CitoplasmaDocumento5 páginasCitoplasmaUri GonzalezAún no hay calificaciones
- El Citoplasma y Sus OrganelosDocumento4 páginasEl Citoplasma y Sus OrganelosPT DtupAún no hay calificaciones
- Sistema EndomembranasDocumento25 páginasSistema EndomembranasGil Ruano100% (1)
- Los Ribosomas - BiologiaDocumento7 páginasLos Ribosomas - BiologiaMag SosaAún no hay calificaciones
- Manual de FisiologìaDocumento120 páginasManual de Fisiologìaisabel reyesAún no hay calificaciones
- Celula AnimalDocumento11 páginasCelula AnimalDania FloresAún no hay calificaciones
- Bloque 2 (II) Componentes de La Célula Eucariota.Documento33 páginasBloque 2 (II) Componentes de La Célula Eucariota.HIDRANTE COLUMNA SECAAún no hay calificaciones
- Resumen de Fisiologia Celular.Documento12 páginasResumen de Fisiologia Celular.Patricia De JesúsAún no hay calificaciones
- Aparato de Golgi o DictiosomaDocumento9 páginasAparato de Golgi o DictiosomaAnonymous QNfesIQxAún no hay calificaciones
- Ciclos de La MateriaDocumento5 páginasCiclos de La MateriaLuz Ang�lica Sandoval S�nchezAún no hay calificaciones
- EPITELIODocumento5 páginasEPITELIOLuz Ang�lica Sandoval S�nchezAún no hay calificaciones
- La Biosfera y Sus ComponentesDocumento3 páginasLa Biosfera y Sus ComponentesLuz Ang�lica Sandoval S�nchezAún no hay calificaciones
- Biomas Acuáticos y TerrestresDocumento5 páginasBiomas Acuáticos y TerrestresLuz Ang�lica Sandoval S�nchezAún no hay calificaciones
- Unidad Educativa Liceo Cristiano de Guayaquil: Taller 1 de Biologia: TemaDocumento12 páginasUnidad Educativa Liceo Cristiano de Guayaquil: Taller 1 de Biologia: TemaDeisy JTigreAún no hay calificaciones
- Taller de BiologíaDocumento3 páginasTaller de BiologíaMARTIN FELIPE CASAS RAMOSAún no hay calificaciones
- Qué Es La FisiologíaDocumento8 páginasQué Es La FisiologíaelenaAún no hay calificaciones
- Diapositivas Lisosomas y PeroxisomasDocumento5 páginasDiapositivas Lisosomas y PeroxisomasHector SuarezAún no hay calificaciones
- Estructura Del Citoplasma para Cuarto Grado de SecundariaDocumento6 páginasEstructura Del Citoplasma para Cuarto Grado de Secundarialeo apazaAún no hay calificaciones
- BiologiaDocumento17 páginasBiologiaJeancarlos Mendoza VillarroelAún no hay calificaciones
- NucleoDocumento28 páginasNucleoRoberto ItzepAún no hay calificaciones
- Biología Orgamelos CelularesDocumento5 páginasBiología Orgamelos CelularesFujoshi YomeilyAún no hay calificaciones
- Lectura Complemento-Vesículas Recubiertas de COPDocumento8 páginasLectura Complemento-Vesículas Recubiertas de COPEvelyn FloresAún no hay calificaciones
- Amelogénesis y Dentinogénesis - Versión PreliminarDocumento21 páginasAmelogénesis y Dentinogénesis - Versión PreliminarAndres GonzalezAún no hay calificaciones
- Organoides Celulares ResumenDocumento24 páginasOrganoides Celulares ResumenAlejandro BrodersAún no hay calificaciones
- Aparato de GolgiDocumento34 páginasAparato de GolgiMoisés J. Duran CanevaroAún no hay calificaciones
- Medicina CelularDocumento439 páginasMedicina CelularLuis Jorge Urrea MoralesAún no hay calificaciones
- Segundo Parcial Biologia General 2021 P1Documento175 páginasSegundo Parcial Biologia General 2021 P1Monica RodriguezAún no hay calificaciones
- Dossier Introducción A La Biología CelularDocumento7 páginasDossier Introducción A La Biología CelularBarbie KenAún no hay calificaciones
- Cuadros Resumen Célula y OrgánelosDocumento17 páginasCuadros Resumen Célula y OrgánelosJORGE ENRIQUE MEDINA CABRERAAún no hay calificaciones
- Evaluacion Final - Escenario 8 - PRIMER BLOQUE-TEORICO - BIOLOGIA HUMANA - (GRUPO B01)Documento22 páginasEvaluacion Final - Escenario 8 - PRIMER BLOQUE-TEORICO - BIOLOGIA HUMANA - (GRUPO B01)Sandra RamirezAún no hay calificaciones
- Guias de Laboratorio BTA B40010 1-2017Documento33 páginasGuias de Laboratorio BTA B40010 1-2017Marlys CamposAún no hay calificaciones
- Tabla Orgánulos Hwysgqh-1Documento1 páginaTabla Orgánulos Hwysgqh-1ruthAún no hay calificaciones
- La CelulaDocumento2 páginasLa CelulaIsabella Marquez PalominoAún no hay calificaciones
- Lectura Complementaria Fisiología 1 - Morfofunción 1 PDFDocumento47 páginasLectura Complementaria Fisiología 1 - Morfofunción 1 PDFCristian0% (1)
- Diccionario Ilustrado - Botanica EverestDocumento108 páginasDiccionario Ilustrado - Botanica EverestFede Jdb100% (2)
- Reporte 2 PlantasDocumento5 páginasReporte 2 PlantasAngy MarquezAún no hay calificaciones
- Unidad 06 Sistema de EndomembranasDocumento9 páginasUnidad 06 Sistema de EndomembranasjvlpAún no hay calificaciones
- Partes de La CelulaDocumento1 páginaPartes de La CelulaOsmandy VeraAún no hay calificaciones
- María Alejandra Correa Pons - Actividades CélulaDocumento15 páginasMaría Alejandra Correa Pons - Actividades CélulaMaria CorreaAún no hay calificaciones
- Banco de Preguntas de CienciasDocumento4 páginasBanco de Preguntas de CienciasmariaAún no hay calificaciones
- Citologia Celula Eucariota Citoplasma NucleoDocumento4 páginasCitologia Celula Eucariota Citoplasma NucleoAndre Fernandez0% (1)
- Banco de Preguntas de Biologia.Documento85 páginasBanco de Preguntas de Biologia.Cristian MamaniAún no hay calificaciones
- Guía N°1 La Célula Animal y Vegetal 8° BASICODocumento3 páginasGuía N°1 La Célula Animal y Vegetal 8° BASICOALM - GAEL AQUILES BUSTOSAún no hay calificaciones