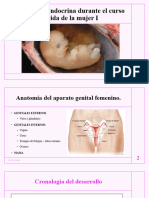
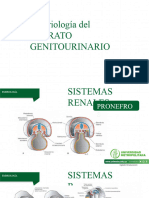
También podría gustarte
- Embriología Del Aparato ReproductorDocumento83 páginasEmbriología Del Aparato ReproductorEdgardo Leal VillarroelAún no hay calificaciones
- S GenitalDocumento16 páginasS GenitalganardineropersodeanaAún no hay calificaciones
- Embrio FemDocumento3 páginasEmbrio FemValenAún no hay calificaciones
- Clase 4 Ginecología IDocumento29 páginasClase 4 Ginecología Idayana cruzAún no hay calificaciones
- Cuadro Comparativo Del Desarrollo de La GónadaDocumento14 páginasCuadro Comparativo Del Desarrollo de La GónadaMiguel Angel Parra GarcíaAún no hay calificaciones
- Gónadas, Gametos y GametogenesisDocumento56 páginasGónadas, Gametos y GametogenesisJonathan Montesinos Delgado100% (1)
- Semana 2, GametogenesisDocumento67 páginasSemana 2, GametogenesisMichelle MorenoAún no hay calificaciones
- s.genital Resumen CapDocumento13 páginass.genital Resumen Capalejahernandez3120Aún no hay calificaciones
- Sistema Reproductor CompletoDocumento75 páginasSistema Reproductor CompletoGeraldinne SuarezAún no hay calificaciones
- Embriología de los órganos reproductoresDocumento49 páginasEmbriología de los órganos reproductoresDanielaAnguloAún no hay calificaciones
- Aparato Reproductor Masculino 2023Documento17 páginasAparato Reproductor Masculino 2023annabella del vecchioAún no hay calificaciones
- Sexualidad, Genero y ReproduccionDocumento163 páginasSexualidad, Genero y ReproduccionNoelia MontenegroAún no hay calificaciones
- Embriologia IIDocumento34 páginasEmbriologia IIMaryRuizAún no hay calificaciones
- Determinación Sexual Mujer - Documentos de Google PDFDocumento14 páginasDeterminación Sexual Mujer - Documentos de Google PDFOctavio BorriAún no hay calificaciones
- Embriologia AnimalDocumento6 páginasEmbriologia Animalmaría belén pacheco reyesAún no hay calificaciones
- Resumen Desarrollo y GenéticaDocumento59 páginasResumen Desarrollo y GenéticaSantehhhAún no hay calificaciones
- La Saga de La Linea GerminalDocumento54 páginasLa Saga de La Linea GerminalCarlos Julio Alvarez BetinAún no hay calificaciones
- Clase 12 - Sistema ReproductorDocumento122 páginasClase 12 - Sistema ReproductorElizabeth GarridoAún no hay calificaciones
- Desarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexoDocumento19 páginasDesarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexojuanAún no hay calificaciones
- Gonadas EmbriologiaDocumento1 páginaGonadas EmbriologiaMariana AltamiranoAún no hay calificaciones
- Resumen de Aparato Reproductor Masculino DesiDocumento7 páginasResumen de Aparato Reproductor Masculino DesiDaya GarciaAún no hay calificaciones
- Capitulo 18 Embriogenesis Aparato Genital 2017Documento16 páginasCapitulo 18 Embriogenesis Aparato Genital 2017sandraAún no hay calificaciones
- Modelo Apuntes Embriologia BIRDocumento7 páginasModelo Apuntes Embriologia BIRMaria del Rosario Aguilar MayoAún no hay calificaciones
- La Saga de La Linea GerminalDocumento54 páginasLa Saga de La Linea GerminalEgresados BilologiaAún no hay calificaciones
- 1Documento5 páginas1Mikaela lAún no hay calificaciones
- Desarrollo embrionario del aparato reproductor femeninoDocumento30 páginasDesarrollo embrionario del aparato reproductor femeninoAlejandro CruzAún no hay calificaciones
- UrogenitalDocumento40 páginasUrogenitalElianneAún no hay calificaciones
- Resumen - Fisio Reproductiva (Berne y Levy)Documento20 páginasResumen - Fisio Reproductiva (Berne y Levy)Yara MartinezAún no hay calificaciones
- Conceptos básicos de embriología enDocumento53 páginasConceptos básicos de embriología enMichelle La OrgaAún no hay calificaciones
- Embriología genital masculina y femeninaDocumento17 páginasEmbriología genital masculina y femeninaAna Cristina Sanchez ZangAún no hay calificaciones
- EmbriologíaDocumento230 páginasEmbriologíaMelissa PomaAún no hay calificaciones
- Gametogenesis, Fecundacion y Embriogenesis 1Documento13 páginasGametogenesis, Fecundacion y Embriogenesis 1Andreina GarciaAún no hay calificaciones
- Gametogénesis y meiosis: La formación de gametosDocumento7 páginasGametogénesis y meiosis: La formación de gametosHeidy Juliana Ramírez GómezAún no hay calificaciones
- ESpermatogenesisDocumento7 páginasESpermatogenesismagali cruzado caspitoAún no hay calificaciones
- Aparato Genital MasculinoDocumento30 páginasAparato Genital Masculinojames cedeAún no hay calificaciones
- Mitosis, Meiosis, Gametogenesis, GenomaDocumento24 páginasMitosis, Meiosis, Gametogenesis, GenomaEric Abel Villanueva QuintanillaAún no hay calificaciones
- Embriologia Anatomia e Histologia Del Aparato Reproductor MasculinoDocumento59 páginasEmbriologia Anatomia e Histologia Del Aparato Reproductor MasculinoramirezherrerayovanaemiliAún no hay calificaciones
- Informe 2Documento7 páginasInforme 2CHAVARRI REGALADO STEPHANIA DE GUADALUPEAún no hay calificaciones
- Resumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital MasculinoDocumento13 páginasResumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital Masculinovictoriaetchegoyen123Aún no hay calificaciones
- Presentación 1Documento36 páginasPresentación 1Diego BarrientosAún no hay calificaciones
- Embrilogia Del Sistema Genital Masculino y FemenioDocumento27 páginasEmbrilogia Del Sistema Genital Masculino y FemenioOctavio Villacrez OlascoagaAún no hay calificaciones
- Repaso de EmbriologiaDocumento8 páginasRepaso de EmbriologiaNicole HernandezAún no hay calificaciones
- GAMETOGÉNESISDocumento32 páginasGAMETOGÉNESISDayana PaolaAún no hay calificaciones
- Gametogénesis Masculina y Fecundación 1Documento10 páginasGametogénesis Masculina y Fecundación 1ferlagosmontecinosAún no hay calificaciones
- Desarrolloembrionario PDFDocumento19 páginasDesarrolloembrionario PDFAlejandro DávilaAún no hay calificaciones
- Las Células Germinales Primordiales Migran Por El Mesenterio Dorsal Desde El Saco Vitelino Hacia Las Crestas GenitalesDocumento1 páginaLas Células Germinales Primordiales Migran Por El Mesenterio Dorsal Desde El Saco Vitelino Hacia Las Crestas GenitalesHannie AbudAún no hay calificaciones
- Embriología de GenitalesDocumento7 páginasEmbriología de GenitalesViviana PalaciosAún no hay calificaciones
- EMBRIOLOGIA DEL SISTEMA GENITOURINARIODocumento22 páginasEMBRIOLOGIA DEL SISTEMA GENITOURINARIOJennifer CampoAún no hay calificaciones
- Biologia de La Concepcion Aparato Reproductor (Autoguardado)Documento16 páginasBiologia de La Concepcion Aparato Reproductor (Autoguardado)Eliana Velasco100% (1)
- GametogenesisDocumento11 páginasGametogenesisVanessa Rodriguez OlmosAún no hay calificaciones
- Desarrollo de Las Gonadas Masculinas y FemeninasDocumento3 páginasDesarrollo de Las Gonadas Masculinas y FemeninasAle AntonioAún no hay calificaciones
- 2 GametogenesisDocumento61 páginas2 GametogenesisOlivia Berenisse Corona EspinozaAún no hay calificaciones
- GametogenesisDocumento9 páginasGametogenesismiguel minierAún no hay calificaciones
- A Espermatogenesis. Arteaga M. Modif.Documento62 páginasA Espermatogenesis. Arteaga M. Modif.Brigith CruzAún no hay calificaciones
- Embriología de Las GonadasDocumento16 páginasEmbriología de Las GonadasShirley GarciaAún no hay calificaciones
- Repro Animal U4 - 3 EspermatogénesisDocumento25 páginasRepro Animal U4 - 3 EspermatogénesisAnita Yael Molina EstelaAún no hay calificaciones
- Guia Morfologia 1Documento11 páginasGuia Morfologia 1Ana murilloAún no hay calificaciones
- Espermatogenesis y EspermiogenesisDocumento4 páginasEspermatogenesis y EspermiogenesisMónica Sinaí Macías HernándezAún no hay calificaciones
- Repaso de Embriologia CTO DR Carlos Gonzales MedinaDocumento216 páginasRepaso de Embriologia CTO DR Carlos Gonzales MedinaCarlos GonzalesAún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Clasificación y definición de ortodoncia preventiva, interceptiva y correctivaDocumento1 páginaClasificación y definición de ortodoncia preventiva, interceptiva y correctivaDaniela GonzálezAún no hay calificaciones
- VARIABLESDocumento4 páginasVARIABLESDaniela GonzálezAún no hay calificaciones
- E. coli 40Documento5 páginasE. coli 40Daniela GonzálezAún no hay calificaciones
- Interrelaciones de Las Funciones Respiratorias Masticatorias, Deglución y FonaciónDocumento1 páginaInterrelaciones de Las Funciones Respiratorias Masticatorias, Deglución y FonaciónDaniela GonzálezAún no hay calificaciones
- Caries de La Infancia Temprana SeveraDocumento16 páginasCaries de La Infancia Temprana SeveraDaniela GonzálezAún no hay calificaciones
- Tipos de Estudio en InvestigaciónDocumento2 páginasTipos de Estudio en InvestigaciónDaniela GonzálezAún no hay calificaciones
- Trazado Cefalométrico FinalizadoDocumento1 páginaTrazado Cefalométrico FinalizadoDaniela GonzálezAún no hay calificaciones
- Expediente ClínicoDocumento2 páginasExpediente ClínicoDaniela GonzálezAún no hay calificaciones
- Ortodoncia PreventivaDocumento3 páginasOrtodoncia PreventivaDaniela GonzálezAún no hay calificaciones
- Técnicas de Conformado y EndurecimientoDocumento2 páginasTécnicas de Conformado y EndurecimientoAlan JadirtAún no hay calificaciones
- Coronilla A La Divina MisericordiaDocumento3 páginasCoronilla A La Divina MisericordiaJorge Ignacio Tapia BaldrichAún no hay calificaciones
- Formato de RiesgoDocumento11 páginasFormato de RiesgoJuan Carlos Huaman HuamaniAún no hay calificaciones
- Catálogo Kolorines 2010Documento61 páginasCatálogo Kolorines 2010UzielAún no hay calificaciones
- Consepto de Actividad Fisica Con Sus Propias Palabras VivianaDocumento4 páginasConsepto de Actividad Fisica Con Sus Propias Palabras VivianaNory MariinAún no hay calificaciones
- Glosario Ovino - Caprino 2 CorteDocumento13 páginasGlosario Ovino - Caprino 2 CorteCarol Daniela GarzonAún no hay calificaciones
- Con La Segunda Bandera en El Frente de Aragon by Francisco Cavero Y Cavero (Z-Lib - Org) .fb2Documento92 páginasCon La Segunda Bandera en El Frente de Aragon by Francisco Cavero Y Cavero (Z-Lib - Org) .fb2Felipe QuintoAún no hay calificaciones
- Matematica 3Documento7 páginasMatematica 3Efrain Correa VelascoAún no hay calificaciones
- Evaluacion Fono en NeoDocumento5 páginasEvaluacion Fono en NeoFabiola Valdebenito PezoAún no hay calificaciones
- PDF 2, SEMANA 2 Informe CualitativoDocumento30 páginasPDF 2, SEMANA 2 Informe CualitativoBrenda Pérez BurtonAún no hay calificaciones
- Aula Virtual Semana 12 'La Agrupación EspacioDocumento10 páginasAula Virtual Semana 12 'La Agrupación EspacioFabiola CariapazaAún no hay calificaciones
- Diccionario de Terminos PiagetianosDocumento18 páginasDiccionario de Terminos PiagetianosLuis Eduardo Salas Rios100% (2)
- Evaluacion Diagnostica de 3ro.Documento3 páginasEvaluacion Diagnostica de 3ro.Agueda Arelis Acosta CastilloAún no hay calificaciones
- Maxi RestDocumento20 páginasMaxi RestGA100% (1)
- Codoñer, Carmen (Ed.) - Historia de La Literatura Latina Catulo y HoracioDocumento27 páginasCodoñer, Carmen (Ed.) - Historia de La Literatura Latina Catulo y Horaciogabriela curcuyAún no hay calificaciones
- Orica (1) HDJWJDocumento26 páginasOrica (1) HDJWJFelipe BertolottoAún no hay calificaciones
- Preguntas de Autoevaluación Capítulo 24Documento4 páginasPreguntas de Autoevaluación Capítulo 24oscar26108750% (2)
- Alteraciones del Yo: Dimensiones y psicopatología enDocumento17 páginasAlteraciones del Yo: Dimensiones y psicopatología enJorge Alberto Mestiza GarcíaAún no hay calificaciones
- Acuerdo entre el ISFD N° 1 y ATEN para acciones de formación docenteDocumento4 páginasAcuerdo entre el ISFD N° 1 y ATEN para acciones de formación docenteLuis Angel SolavagioneAún no hay calificaciones
- Refrigeracion Domestica TerminadoDocumento5 páginasRefrigeracion Domestica TerminadoLucas Andres Yanac DurandAún no hay calificaciones
- Copetro Es Una Planta Perteneciente A La Empresa Oxbow Carbon LCCDocumento6 páginasCopetro Es Una Planta Perteneciente A La Empresa Oxbow Carbon LCCLucas VázquezAún no hay calificaciones
- Ecuación DiscriminanteDocumento8 páginasEcuación DiscriminanteNicol CahuapasAún no hay calificaciones
- TOC - ClozapinaDocumento3 páginasTOC - ClozapinaPablo MenendezAún no hay calificaciones
- Sociologia Con Isse Moyano - Resumen (Full Permission)Documento18 páginasSociologia Con Isse Moyano - Resumen (Full Permission)Esga Myrina MiaAún no hay calificaciones
- FERRERO DIEZ CANSECO, A., Las Formas Especiales de Sociedad Anónima en La Nueva LGSDocumento17 páginasFERRERO DIEZ CANSECO, A., Las Formas Especiales de Sociedad Anónima en La Nueva LGSYakelyn SussanAún no hay calificaciones
- Sifón Proyecto Olmos TinajonesDocumento16 páginasSifón Proyecto Olmos TinajonesArturo Rafael Olguin NoriegaAún no hay calificaciones
- G23-Objetivos, Enfoques y Elementos Del CurriculumDocumento9 páginasG23-Objetivos, Enfoques y Elementos Del CurriculumJonathan ZuñigaAún no hay calificaciones
- Ejercicios EstadisticaDocumento6 páginasEjercicios EstadisticaLuarSoft20% (1)
- Tarea 4 Pensamiento Logico MatematicoDocumento9 páginasTarea 4 Pensamiento Logico MatematicoMerlyciita RozoAún no hay calificaciones