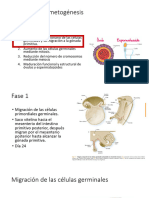
También podría gustarte
- La Saga de La Linea GerminalDocumento54 páginasLa Saga de La Linea GerminalCarlos Julio Alvarez BetinAún no hay calificaciones
- GametogenesisDocumento11 páginasGametogenesisVanessa Rodriguez OlmosAún no hay calificaciones
- ESpermatogenesisDocumento7 páginasESpermatogenesismagali cruzado caspitoAún no hay calificaciones
- Biologia de La Concepcion Aparato Reproductor (Autoguardado)Documento16 páginasBiologia de La Concepcion Aparato Reproductor (Autoguardado)Eliana Velasco100% (1)
- Repaso de EmbriologiaDocumento8 páginasRepaso de EmbriologiaNicole HernandezAún no hay calificaciones
- Gamtogénesis by @med - TropolisDocumento8 páginasGamtogénesis by @med - TropolisJean SalgadoAún no hay calificaciones
- GAMETOGENESISDocumento6 páginasGAMETOGENESISCamilaAún no hay calificaciones
- EspermatogénesisDocumento5 páginasEspermatogénesisXavier GuillenAún no hay calificaciones
- EspermatogénesisDocumento3 páginasEspermatogénesisSteveAún no hay calificaciones
- Modelo Apuntes Embriologia BIRDocumento7 páginasModelo Apuntes Embriologia BIRMaria del Rosario Aguilar MayoAún no hay calificaciones
- GametogenesisDocumento50 páginasGametogenesisEmiliano CardenasAún no hay calificaciones
- Capitulo 2 - Langman - GametogénesisDocumento5 páginasCapitulo 2 - Langman - Gametogénesismaria jose sanchez gilAún no hay calificaciones
- Celulas Germinales PrimordialesDocumento9 páginasCelulas Germinales PrimordialesDora Urbano100% (1)
- 01 Gametogénesis y FecundaciónDocumento16 páginas01 Gametogénesis y FecundaciónAdolfo Antonio Ríos AlcortaAún no hay calificaciones
- Embriología - Gametogénesis y Ciclo OváricoDocumento7 páginasEmbriología - Gametogénesis y Ciclo OváricoHeidy Juliana Ramírez GómezAún no hay calificaciones
- Determinación Sexual Mujer - Documentos de Google PDFDocumento14 páginasDeterminación Sexual Mujer - Documentos de Google PDFOctavio BorriAún no hay calificaciones
- Resumen Desarrollo y GenéticaDocumento59 páginasResumen Desarrollo y GenéticaSantehhhAún no hay calificaciones
- GametogénesisDocumento28 páginasGametogénesisSofia PantojaAún no hay calificaciones
- R e S U M e N Ciclo OváricoDocumento3 páginasR e S U M e N Ciclo OváricoMaria Fernanda Pizarro VasquezAún no hay calificaciones
- GAMETOGÉNESISDocumento32 páginasGAMETOGÉNESISDayana PaolaAún no hay calificaciones
- Tema 4Documento8 páginasTema 4Alessandra CancemiAún no hay calificaciones
- EspermatogenesisDocumento5 páginasEspermatogenesisJuan Jose Garcia MoralesAún no hay calificaciones
- GAMETOGÉNEIS - EspermatogénesisDocumento22 páginasGAMETOGÉNEIS - EspermatogénesisAsriel CoulterAún no hay calificaciones
- EMI MonografiaDocumento25 páginasEMI MonografiaAlejandro Mendoza SarmientoAún no hay calificaciones
- GametogenesisDocumento9 páginasGametogenesismiguel minierAún no hay calificaciones
- FECUNDACIONDocumento37 páginasFECUNDACIONDian OrtizAún no hay calificaciones
- Espermatogénesis en AnimalesDocumento3 páginasEspermatogénesis en AnimalesFernanda Overstreet0% (1)
- GametogénesisDocumento27 páginasGametogénesisSergio Alejandro IbarraAún no hay calificaciones
- Trabajo de EmbriologiaDocumento9 páginasTrabajo de EmbriologiaJosé CastroAún no hay calificaciones
- Hormonas Sexuales MasculinasDocumento25 páginasHormonas Sexuales MasculinasJoselyn GonzalezAún no hay calificaciones
- Gametogenesis AlumnosDocumento48 páginasGametogenesis AlumnospinedobautistaedAún no hay calificaciones
- Guia Gametogenesis, Fecundacion y 1ra SemanaDocumento5 páginasGuia Gametogenesis, Fecundacion y 1ra SemanaeldoctopuntocomAún no hay calificaciones
- Ao 10Documento5 páginasAo 10Maria TorresAún no hay calificaciones
- Embriologia2 PDFDocumento47 páginasEmbriologia2 PDFFlavia Cristina SilvaAún no hay calificaciones
- Tema 29 Nociones de La EmbriologíaDocumento7 páginasTema 29 Nociones de La EmbriologíaAURILETHAún no hay calificaciones
- Desarrollo EmbrionarioDocumento23 páginasDesarrollo EmbrionarioCamilo Carné Candil67% (3)
- Presentación 1Documento36 páginasPresentación 1Diego BarrientosAún no hay calificaciones
- E1 GametogénesisDocumento7 páginasE1 GametogénesisKaryme PérezAún no hay calificaciones
- EMBRIOLOGÍA 4 GAMETOGÉNESIS, FECUNDACIÓN E IMPLANTACIÓN (1) (1) .OdpDocumento68 páginasEMBRIOLOGÍA 4 GAMETOGÉNESIS, FECUNDACIÓN E IMPLANTACIÓN (1) (1) .OdpNaydelin BravoAún no hay calificaciones
- Game To GenesisDocumento36 páginasGame To Genesismaria ines rico zeferinoAún no hay calificaciones
- GametogenesisDocumento41 páginasGametogenesishiroshy555Aún no hay calificaciones
- Espermatogenesis - TrabajoDocumento12 páginasEspermatogenesis - TrabajoJanella Tineo MedinaAún no hay calificaciones
- Gametogenesis 2Documento24 páginasGametogenesis 2Jherson TerrazasAún no hay calificaciones
- EmbriologíaDocumento230 páginasEmbriologíaMelissa PomaAún no hay calificaciones
- Espermatología ForenseDocumento13 páginasEspermatología ForenseYahir Esau Gutierrez FloresAún no hay calificaciones
- EMBRIOLOGÌA - Documentos de GoogleDocumento8 páginasEMBRIOLOGÌA - Documentos de GoogleLeslie SánchezAún no hay calificaciones
- Repro Animal U4 - 3 EspermatogénesisDocumento25 páginasRepro Animal U4 - 3 EspermatogénesisAnita Yael Molina EstelaAún no hay calificaciones
- GAMETOGENESISDocumento84 páginasGAMETOGENESISMery de Aguilar100% (1)
- Sexualidad, Genero y ReproduccionDocumento163 páginasSexualidad, Genero y ReproduccionNoelia MontenegroAún no hay calificaciones
- 1.introduccion A La Embriologia y Gametogenesis-1Documento26 páginas1.introduccion A La Embriologia y Gametogenesis-1Bryan Amaru GonzálezAún no hay calificaciones
- 2 Gametogenesis, Fecundacion y ImplantacionDocumento37 páginas2 Gametogenesis, Fecundacion y Implantacionyasser08 Hernandez100% (1)
- Gametogénesis y FecundaciónDocumento15 páginasGametogénesis y FecundaciónM.E. PirelaAún no hay calificaciones
- EspermatogenesisDocumento5 páginasEspermatogenesisDiana Jimenez MartinezAún no hay calificaciones
- AO #7. Desarrollo Pre-Embrionario..Documento12 páginasAO #7. Desarrollo Pre-Embrionario..Andrea MedinaAún no hay calificaciones
- Guia Morfologia 1Documento11 páginasGuia Morfologia 1Ana murilloAún no hay calificaciones
- Resumen GametogenesisDocumento3 páginasResumen GametogenesisErik Lopez MamaniAún no hay calificaciones
- Gametognesis BiologiaDocumento15 páginasGametognesis BiologiaJosue RebolledoAún no hay calificaciones
- Sistema ResumidoDocumento24 páginasSistema ResumidoJorge RamirezAún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- 1PM1 - Garces Jurado Jesus - Histologia Testiculo y OvarioDocumento8 páginas1PM1 - Garces Jurado Jesus - Histologia Testiculo y OvarioJesusTauroAún no hay calificaciones
- Planeacion 4°b Semana09Documento9 páginasPlaneacion 4°b Semana09Diaanitha PérezAún no hay calificaciones
- T7 Ciclo Sexual y AnticonceptivosDocumento72 páginasT7 Ciclo Sexual y AnticonceptivosMendez Torres Zaid AlanAún no hay calificaciones
- Anatomía Del Aparato Reproductor Del VerracoDocumento19 páginasAnatomía Del Aparato Reproductor Del VerracoMercedes Areco100% (1)
- 6 Planificacion y CuadernilloDocumento41 páginas6 Planificacion y CuadernilloXimena Vásquez Melgarejo67% (6)
- Unidad 1reproducción AnimalDocumento8 páginasUnidad 1reproducción AnimalAdele FarynaAún no hay calificaciones
- Clase 3 Anatomia Funcional Del MachoDocumento21 páginasClase 3 Anatomia Funcional Del Machojose carlos jimenez huashuayoAún no hay calificaciones
- Sistema Reproductor HumanoDocumento4 páginasSistema Reproductor HumanoSTEFANNY LIZBETH DE LA CRUZ ESPINOZAAún no hay calificaciones
- 20 - Fisiologia de La Reproduccion en El MachoDocumento4 páginas20 - Fisiologia de La Reproduccion en El MachoEmerson Oriel Cedeño González100% (1)
- VaginaDocumento9 páginasVaginaastrosAún no hay calificaciones
- Sem 10 Malformaciones Aparato Genito Urinario UrogenitalDocumento6 páginasSem 10 Malformaciones Aparato Genito Urinario UrogenitalHappier. subsAún no hay calificaciones
- Primera Semana de Desarrollo EmbrionarioDocumento13 páginasPrimera Semana de Desarrollo EmbrionarioLuli CamposAún no hay calificaciones
- Catálogo Digital-Biología 4Documento17 páginasCatálogo Digital-Biología 4Paola Almeida100% (1)
- PY096212 - Reproduccion - Animales - Domesticos - Dosier - EDRA SPDocumento18 páginasPY096212 - Reproduccion - Animales - Domesticos - Dosier - EDRA SPJose E. MartinezAún no hay calificaciones
- Nueva Evidencia Cientifica para El Manejo de La HBP. Tratamiento de Inicio, Cuando y Por Que - Dr. J.M. PalaciosDocumento48 páginasNueva Evidencia Cientifica para El Manejo de La HBP. Tratamiento de Inicio, Cuando y Por Que - Dr. J.M. PalaciosAlexander CastilloAún no hay calificaciones
- VaricoceleDocumento19 páginasVaricoceleSamuel González CandiaAún no hay calificaciones
- Hipogonadismo Secundario Del Adulto - Dr. Christian Safra M.Documento67 páginasHipogonadismo Secundario Del Adulto - Dr. Christian Safra M.ANtonyMVAún no hay calificaciones
- Sistema Reproductor Femenino para Cuarto de PrimariaDocumento4 páginasSistema Reproductor Femenino para Cuarto de PrimariaJhannes Rojas100% (1)
- Pez de AnchanteDocumento21 páginasPez de AnchanteJamyl Javier EchegarayAún no hay calificaciones
- Tablas de Niveles de Testosterona Por Edad (Total y Libre) - Tu SaludDocumento11 páginasTablas de Niveles de Testosterona Por Edad (Total y Libre) - Tu Saludluis sansAún no hay calificaciones
- Mes de MayoDocumento28 páginasMes de MayoMirtha Romero DiazAún no hay calificaciones
- Cooperia SPPDocumento22 páginasCooperia SPPJosé Medina100% (1)
- Sindrome de Ovario Poliquaitico. Angela Viviana SanchezDocumento33 páginasSindrome de Ovario Poliquaitico. Angela Viviana Sanchezmargarita rodriguezAún no hay calificaciones
- Tema 6 La Funcion de ReproduccionDocumento21 páginasTema 6 La Funcion de Reproduccionantonio joseAún no hay calificaciones
- Sistema Reproductor Humano para Sexto de PrimariaDocumento13 páginasSistema Reproductor Humano para Sexto de Primarianeshmy sharomAún no hay calificaciones
- Desarrollo Del Adolescente PDFDocumento6 páginasDesarrollo Del Adolescente PDFsandraesmeraldaAún no hay calificaciones
- Metamorfosis de La PubertadDocumento3 páginasMetamorfosis de La PubertadConstanza ArozaAún no hay calificaciones
- Aparato MasculinoDocumento45 páginasAparato MasculinoDavid Sanchez100% (1)
- 28 ObtenciÓn y Propiedades Del HidrogenoDocumento23 páginas28 ObtenciÓn y Propiedades Del Hidrogenoclaudia vasquezAún no hay calificaciones
- Anatomia Del Aparato Femenino.1GDocumento5 páginasAnatomia Del Aparato Femenino.1GSebastianAún no hay calificaciones