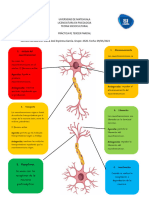
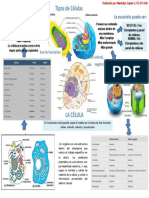
También podría gustarte
- I-4. Diversidad InmunológicaDocumento11 páginasI-4. Diversidad InmunológicaSergiollafeAún no hay calificaciones
- Trabajo de Investigación InmunologíaDocumento7 páginasTrabajo de Investigación Inmunologíacarlina villamarAún no hay calificaciones
- Inducción de La Respuesta InmunitaríaDocumento56 páginasInducción de La Respuesta InmunitaríaOmar PopocaAún no hay calificaciones
- Inmunología EstomatologicaDocumento1 páginaInmunología EstomatologicaDiego jesús Arias MendozaAún no hay calificaciones
- Sistema Inmune y GenéticaDocumento18 páginasSistema Inmune y GenéticaIan Ignacio AlvaradoAún no hay calificaciones
- Resumen Y Guía De Estudio - La Espléndida Curación: Aprovechando Las Defensas Naturales De Tu CuerpoDe EverandResumen Y Guía De Estudio - La Espléndida Curación: Aprovechando Las Defensas Naturales De Tu CuerpoCalificación: 4 de 5 estrellas4/5 (1)
- Teoría de La InmunidadDocumento13 páginasTeoría de La InmunidadLorena Rodríguez GoezAún no hay calificaciones
- Final de InmunologiaDocumento87 páginasFinal de InmunologiaValentina Trujillo RodríguezAún no hay calificaciones
- Definir Los Conceptos de Inmunidad Innata 01Documento17 páginasDefinir Los Conceptos de Inmunidad Innata 01•Maria Mercedes•Aún no hay calificaciones
- Historia de La InmunologíaDocumento13 páginasHistoria de La InmunologíaSsamm RenteriiaAún no hay calificaciones
- Apuntes de InmunoDocumento103 páginasApuntes de InmunoCarolinaAún no hay calificaciones
- Evolución del pensamiento inmunológicoDocumento9 páginasEvolución del pensamiento inmunológicoSilvia DiazAún no hay calificaciones
- Articulo Seleccion ClonalDocumento4 páginasArticulo Seleccion ClonalErik Brayan Erazo NarvaezAún no hay calificaciones
- Aplicaciones Médicas de La InmunogenéticaDocumento7 páginasAplicaciones Médicas de La Inmunogenéticalapp89Aún no hay calificaciones
- Inmuno 1Documento25 páginasInmuno 1LizGresiaAún no hay calificaciones
- Visión Panorámica Del Sistema InmuneDocumento14 páginasVisión Panorámica Del Sistema InmuneCedricDiehlAún no hay calificaciones
- SUEROS Y VACUNAS Sistema InmunitarioDocumento10 páginasSUEROS Y VACUNAS Sistema InmunitarioMary JiménezAún no hay calificaciones
- Muestras Gratis InmunoDocumento45 páginasMuestras Gratis Inmunomicro.fcvunlpAún no hay calificaciones
- Tema 13 Grado FBH 2017Documento21 páginasTema 13 Grado FBH 2017Carlos FelizAún no hay calificaciones
- Inmunologia Medica PDFDocumento106 páginasInmunologia Medica PDFMariaCieloPfAún no hay calificaciones
- Laboratorio 2Documento5 páginasLaboratorio 2ml1374051Aún no hay calificaciones
- HEMATOLOGIA E INMUNOLOGIA UnimarDocumento4 páginasHEMATOLOGIA E INMUNOLOGIA UnimarKATHERINAún no hay calificaciones
- Factores Del ParasitoDocumento16 páginasFactores Del ParasitoantonietaAún no hay calificaciones
- Apuntes Realiza Analisis InmunológicosDocumento15 páginasApuntes Realiza Analisis InmunológicosRuiz Castillo Gabriela NataliaAún no hay calificaciones
- Sistema InmunologicoDocumento10 páginasSistema InmunologicoMariamt RiveroAún no hay calificaciones
- Apuntes Inmuno Completo (1)Documento241 páginasApuntes Inmuno Completo (1)Joaquin IturrarteAún no hay calificaciones
- Articulo de InmunologiaDocumento7 páginasArticulo de InmunologiaNicole Anahi Montes RomeroAún no hay calificaciones
- Práctica 3. Inmunoh Paralelo ADocumento9 páginasPráctica 3. Inmunoh Paralelo AVELASTEGUI PAULINAAún no hay calificaciones
- 7 AÑO - CIENCIAS NATURALES - 2021 - SISTEMA INMUNE - CLASE 3 y 4Documento5 páginas7 AÑO - CIENCIAS NATURALES - 2021 - SISTEMA INMUNE - CLASE 3 y 4Ignacio CatalánAún no hay calificaciones
- Inmuno Resumen-Merged PDFDocumento76 páginasInmuno Resumen-Merged PDFmyriamAún no hay calificaciones
- Tema 1 - Introducción INMUNOLOGIADocumento134 páginasTema 1 - Introducción INMUNOLOGIAHugo Martínez MarínAún no hay calificaciones
- El Sistema InmunológicoDocumento149 páginasEl Sistema Inmunológico17edusAún no hay calificaciones
- Trasplante PreguntasDocumento3 páginasTrasplante PreguntasMaria del carmen ZamudioAún no hay calificaciones
- Inmunidad Innata y AdaptativaDocumento6 páginasInmunidad Innata y AdaptativaDamaris LoorAún no hay calificaciones
- Historia de La Inmunogenética y Células Del SIDocumento27 páginasHistoria de La Inmunogenética y Células Del SIJanieska Dileyne Palacios MartinezAún no hay calificaciones
- 1.la Respuesta InmuneDocumento2 páginas1.la Respuesta InmuneShelonix LongAún no hay calificaciones
- Ensayo. La Respuesta Inmune Innata.Documento7 páginasEnsayo. La Respuesta Inmune Innata.Edgar Omar De La Garza MataAún no hay calificaciones
- PreguntasDocumento18 páginasPreguntasVale Prado VargasAún no hay calificaciones
- El sistema inmune: funciones y componentesDocumento9 páginasEl sistema inmune: funciones y componentesEdgardo Ivan Esparza RicoAún no hay calificaciones
- Filogenia Del Sistema InmuneDocumento11 páginasFilogenia Del Sistema InmuneJacky CarpioAún no hay calificaciones
- InmunologiaDocumento3 páginasInmunologiaJhon GuevaraAún no hay calificaciones
- Inmunologia PediatricaDocumento9 páginasInmunologia PediatricaDaniel VillanuevaAún no hay calificaciones
- InmunologíaDocumento75 páginasInmunologíaPessuto Juan CruzAún no hay calificaciones
- Fundamentos celulares e inmunoterapia actualDocumento52 páginasFundamentos celulares e inmunoterapia actualMikeWasauski ssAún no hay calificaciones
- Revisión Bibliográfica de Inmunidad Mediada Por Las Células.Documento5 páginasRevisión Bibliográfica de Inmunidad Mediada Por Las Células.Jackeline GalarzaAún no hay calificaciones
- DegrabacionesDocumento116 páginasDegrabacionesTania Díez-EderAún no hay calificaciones
- Lección 1Documento42 páginasLección 1Fabiola Rojas JiménezAún no hay calificaciones
- Fagocitosis PDFDocumento7 páginasFagocitosis PDFAna Julia Mero KuffóAún no hay calificaciones
- Tolerancia y Autoinmunidad.Documento6 páginasTolerancia y Autoinmunidad.Sher ValerioAún no hay calificaciones
- INMUNOLOGÍA - Elementos de Respuestas Protectoras Del Organismo AnfitriónDocumento32 páginasINMUNOLOGÍA - Elementos de Respuestas Protectoras Del Organismo AnfitriónRenzo André Casaño100% (7)
- Sistema InmuneDocumento16 páginasSistema InmuneJOSE DUBERLEE TAVARA VEGA100% (1)
- Monografia InmunodiagnosticoDocumento55 páginasMonografia Inmunodiagnosticomapachecon0% (1)
- Sistema InmunologicoDocumento8 páginasSistema Inmunologiconelson samuel quispe itusacaAún no hay calificaciones
- Receptores Tipo Toll PDFDocumento7 páginasReceptores Tipo Toll PDFNiria GamboaAún no hay calificaciones
- Guia InmunologíaDocumento10 páginasGuia InmunologíaFatima JTAún no hay calificaciones
- INMUNODocumento214 páginasINMUNOyexalensmc2107Aún no hay calificaciones
- La Respuesta Inmune. Artículo.Documento8 páginasLa Respuesta Inmune. Artículo.EVANGELINA PÁEZ BUENDÍAAún no hay calificaciones
- 01 Hist&Intr MIDDocumento25 páginas01 Hist&Intr MIDOscar ArshavinAún no hay calificaciones
- 01- INTRODUCCIÓN AL SIST INMUNEDocumento7 páginas01- INTRODUCCIÓN AL SIST INMUNECristAmuert 3Aún no hay calificaciones
- Plag I OoooooDocumento4 páginasPlag I OoooooJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- Abelardo Richard Tesis Bachiller 2018 Part.3 PDFDocumento50 páginasAbelardo Richard Tesis Bachiller 2018 Part.3 PDFMaria Phaola Aguilar FuentesAún no hay calificaciones
- Tesis JALM 2021 Ver2. FinalDocumento117 páginasTesis JALM 2021 Ver2. FinalJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- Ejercicios - Procesos SMAWDocumento2 páginasEjercicios - Procesos SMAWJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- Ingeniería de Materiales Cerámicos 1 - Msc. Rodrigo PereaDocumento3 páginasIngeniería de Materiales Cerámicos 1 - Msc. Rodrigo PereaJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- Calen Covid 2021poDocumento1 páginaCalen Covid 2021poJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- 1er EXAMEN RESUELTO 2021 ADocumento8 páginas1er EXAMEN RESUELTO 2021 AJHON ALEX LOPINTA CCANSAYAAún no hay calificaciones
- TG 2021 PDFDocumento16 páginasTG 2021 PDFJuber Jerson Serrano CervantesAún no hay calificaciones
- Fases de Los NeurotransmisoresDocumento1 páginaFases de Los NeurotransmisoresMaría José EspinosaAún no hay calificaciones
- Transporte Activo y PasivoDocumento2 páginasTransporte Activo y PasivoErika100% (1)
- Preguntas Resueltas de Biologia para Entender Problemas MedicosDocumento120 páginasPreguntas Resueltas de Biologia para Entender Problemas Medicosanghelinebayas58Aún no hay calificaciones
- Listado Libros UCEDocumento34 páginasListado Libros UCEGis CernaAún no hay calificaciones
- ACUAPORINASDocumento41 páginasACUAPORINASLuz CamargoAún no hay calificaciones
- Tema N°11Documento13 páginasTema N°11walter ismael barreto tinedoAún no hay calificaciones
- Examen de Biologia-1Documento2 páginasExamen de Biologia-1Decsy Yojana Almendares NavasAún no hay calificaciones
- Mecanismo de Las ConvulsionesDocumento7 páginasMecanismo de Las ConvulsionesmicaelaAún no hay calificaciones
- Celulas y Moleculas Del Sistema Inmune InnatoDocumento17 páginasCelulas y Moleculas Del Sistema Inmune InnatoDaniela Sadira Rivas MatosAún no hay calificaciones
- Taller Hemograma UNALDocumento4 páginasTaller Hemograma UNALSara ConnorAún no hay calificaciones
- Regulación del receptor de insulina y IRSDocumento2 páginasRegulación del receptor de insulina y IRSKeila RojasAún no hay calificaciones
- Aspectos Generales de Una Hematología CompletaDocumento3 páginasAspectos Generales de Una Hematología CompletanicolAún no hay calificaciones
- Mucosa OralDocumento10 páginasMucosa OralAna María AramayoAún no hay calificaciones
- Anabolismo de Los Carbohidratos PDFDocumento22 páginasAnabolismo de Los Carbohidratos PDFmelyAún no hay calificaciones
- Centriolos y Centros Organizadores HistoDocumento5 páginasCentriolos y Centros Organizadores HistoEstrada Ovando Maria FernandaAún no hay calificaciones
- Tejido EpitelialDocumento71 páginasTejido EpitelialLuis Castañeda ZavaletaAún no hay calificaciones
- Plan de Estudios 2006 - EpmhDocumento4 páginasPlan de Estudios 2006 - EpmhFrancisco Abel SolanoAún no hay calificaciones
- Investigación de Biología MPMDocumento1 páginaInvestigación de Biología MPManakrina1711Aún no hay calificaciones
- Apuntes Del Ingreso Medicina UnicenDocumento52 páginasApuntes Del Ingreso Medicina UnicenRocio del CastilloAún no hay calificaciones
- Clase6 Reproducción Celular 1 1526407194Documento82 páginasClase6 Reproducción Celular 1 1526407194Alberto Delgado EAún no hay calificaciones
- Discucion Bioceel (239Documento18 páginasDiscucion Bioceel (239Ann LeyvaAún no hay calificaciones
- Regeneracion y CicatrizacionDocumento5 páginasRegeneracion y CicatrizacionCristian Flores100% (1)
- La Célula. 6. Peroxisomas, Mitocondrias, Cloroplastos, Gotas de Lípidos. Atlas de Histología Vegetal y AnimalDocumento4 páginasLa Célula. 6. Peroxisomas, Mitocondrias, Cloroplastos, Gotas de Lípidos. Atlas de Histología Vegetal y AnimalWILMA GUADALUPE CASTRO PALMAAún no hay calificaciones
- Prueba de Rally Ciencias GiratoriasDocumento6 páginasPrueba de Rally Ciencias GiratoriasAdriana VergelAún no hay calificaciones
- Introduccion A La BioquimicaDocumento3 páginasIntroduccion A La BioquimicaGabriela LMAún no hay calificaciones
- Libro de Apuntes Bioquímica VegetalDocumento89 páginasLibro de Apuntes Bioquímica VegetalIvi VaughnAún no hay calificaciones
- La Gametogénesis Es La Formación de Gametos Por Medio de La MeiosisDocumento2 páginasLa Gametogénesis Es La Formación de Gametos Por Medio de La MeiosisNanosinx Kress UnityAún no hay calificaciones
- Metabolismo de Las ProteinasDocumento22 páginasMetabolismo de Las ProteinasLizanna Zorrilla CeballosAún no hay calificaciones
- Clase 8 Anabolismo y CatabolismoDocumento13 páginasClase 8 Anabolismo y CatabolismoNinoAún no hay calificaciones
- Mir 01 1718 Preguntastestdeclase Ig 2VDocumento15 páginasMir 01 1718 Preguntastestdeclase Ig 2VLucas OrtizAún no hay calificaciones