También podría gustarte
- Exp. Sist. LinfaticoDocumento7 páginasExp. Sist. Linfaticomanuel meza sotoAún no hay calificaciones
- 19 Sistema LinfaticoDocumento2 páginas19 Sistema LinfaticoAngela Valenzuela VaqueroAún no hay calificaciones
- La Linfa y Su RecorridoDocumento15 páginasLa Linfa y Su RecorridoJhonatan CayllahuaAún no hay calificaciones
- Grupo 5 - Tejido Conectivo Especializado LinfáticoDocumento37 páginasGrupo 5 - Tejido Conectivo Especializado LinfáticoCami Liss0% (1)
- Sistema Linfático, AnatomíaDocumento9 páginasSistema Linfático, AnatomíaNatalia MaldonadoAún no hay calificaciones
- JJJJJJJJJJJJJJDocumento37 páginasJJJJJJJJJJJJJJdulceAún no hay calificaciones
- Sistema LinfáticoDocumento7 páginasSistema LinfáticoDiego Ferrer MonteroAún no hay calificaciones
- Sis LinfaticoDocumento23 páginasSis LinfaticoCitlaliAún no hay calificaciones
- Sistema LinfáticoDocumento11 páginasSistema LinfáticoArlinis ChávezAún no hay calificaciones
- Sistema Linfático CIENCIADocumento9 páginasSistema Linfático CIENCIARamona PinoAún no hay calificaciones
- CaraDocumento18 páginasCaraNicolas Ortiz RojasAún no hay calificaciones
- EsfenoidesDocumento13 páginasEsfenoidesEdson DeschanelAún no hay calificaciones
- Protocolo de Ventas y Servicio Al ClienteDocumento26 páginasProtocolo de Ventas y Servicio Al ClienteRecursos humanos Depco Autopartes100% (1)
- Sistema LinfaticoDocumento11 páginasSistema Linfaticoalexa espinalAún no hay calificaciones
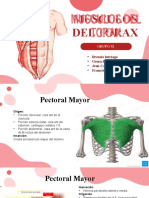
- Expo Musculos Torax 1Documento28 páginasExpo Musculos Torax 1BRENDA INTRIAGO100% (1)
- Músculos, Inervación e Irrigación Del Miembro Superior: Grupo 1Documento62 páginasMúsculos, Inervación e Irrigación Del Miembro Superior: Grupo 1DANIELA LISSET CHINGUEL PALACIOS100% (1)
- Cintura EscapularDocumento96 páginasCintura EscapularAmerissis DowneyAún no hay calificaciones
- Musculos Del Miembro Superior Gray Anatomia para Estudiantes CompressDocumento8 páginasMusculos Del Miembro Superior Gray Anatomia para Estudiantes CompressLaura Matos100% (1)
- Órganos y Tejidos Del Sistema Inmunitari3Documento8 páginasÓrganos y Tejidos Del Sistema Inmunitari3Odalys GarcíaAún no hay calificaciones
- Terminología MédicaDocumento8 páginasTerminología MédicakailaAún no hay calificaciones
- Páginas desdeGUIA RAPIDA M50Documento17 páginasPáginas desdeGUIA RAPIDA M50INTERNET MAFE100% (1)
- Grupo Externo de Musculos Del HombroDocumento14 páginasGrupo Externo de Musculos Del HombroHitler Ivan Chacon QuispeAún no hay calificaciones
- Musculos de La Cara, Cuello y PectoralesDocumento40 páginasMusculos de La Cara, Cuello y PectoralesDesiree Pantaleon AntonAún no hay calificaciones
- Terminologìa QuirùrgicaDocumento6 páginasTerminologìa QuirùrgicaOlga Lucia Duque Quijano100% (1)
- Indicadores Evolutivos en Niños de 9 AñosDocumento2 páginasIndicadores Evolutivos en Niños de 9 AñosalejandraAún no hay calificaciones
- Craneo y Macizo Facial 2012Documento101 páginasCraneo y Macizo Facial 2012juancayalaAún no hay calificaciones
- Musculos Cabeza y CuelloDocumento14 páginasMusculos Cabeza y Cuelloluisa giraldoAún no hay calificaciones
- Drenaje LinfáticoDocumento5 páginasDrenaje Linfáticomexico_lindo754175Aún no hay calificaciones
- Musculos de Cuello y Parte Posterior Del ToraxDocumento38 páginasMusculos de Cuello y Parte Posterior Del ToraxSaulUrquijo100% (1)
- Síndrome Túnel CarpianoDocumento13 páginasSíndrome Túnel CarpianoHospital Río NegroAún no hay calificaciones
- Acne 2 PDFDocumento21 páginasAcne 2 PDFNadia ReineroAún no hay calificaciones
- MELANOGENESISDocumento2 páginasMELANOGENESISprofesorasaraAún no hay calificaciones
- Sistema TegumentarioDocumento9 páginasSistema TegumentarioPatricia Maturana SepulvedaAún no hay calificaciones
- Úlceras de Origen Circulatorio en Miembros Inferiores: IlustradaDocumento57 páginasÚlceras de Origen Circulatorio en Miembros Inferiores: IlustradaCarol VillarAún no hay calificaciones
- Musculos de La Cabeza Cara y CuelloDocumento35 páginasMusculos de La Cabeza Cara y CuelloAlexander OrbeAún no hay calificaciones
- El Drenaje Linfático ManualDocumento8 páginasEl Drenaje Linfático ManualEbert Dante CAún no hay calificaciones
- PelvisDocumento12 páginasPelvisGreidys PadillaAún no hay calificaciones
- Enfermedad Venosa CronicaDocumento14 páginasEnfermedad Venosa CronicaDelfina Zambrano100% (1)
- Escleroterapia Jessmar Spa SpaDocumento25 páginasEscleroterapia Jessmar Spa Spabolesito100% (1)
- CONSENSO SOBRE ULCERAS VASCULARES y PIE DIABETICO Asociacion Española de Enfermeria Vascular PDF GratisDocumento140 páginasCONSENSO SOBRE ULCERAS VASCULARES y PIE DIABETICO Asociacion Española de Enfermeria Vascular PDF GratisDaniel Pérez Silva100% (2)
- Cuestionario Drenaje LinfáticoDocumento18 páginasCuestionario Drenaje LinfáticoAdrianaAún no hay calificaciones
- Velázquez Salinas Dennys - 5HM5 - EF LinfáticoDocumento7 páginasVelázquez Salinas Dennys - 5HM5 - EF LinfáticoVelázquez Salinas DennysAún no hay calificaciones
- Músculos Faciales y MasticadoresDocumento7 páginasMúsculos Faciales y MasticadoresJaviera FuentealbaAún no hay calificaciones
- Anatomia Maqueta Musculos Huesos y ArticulacionesDocumento27 páginasAnatomia Maqueta Musculos Huesos y ArticulacionesClaudia D. AldabaAún no hay calificaciones
- Musculos Expresión FacialDocumento4 páginasMusculos Expresión FacialBarrera Soto Luis AngelAún no hay calificaciones
- 6ta Clase Neuro Tronco Encefalico DR CorreaDocumento53 páginas6ta Clase Neuro Tronco Encefalico DR CorreaAlicia López Gomero100% (1)
- Drenaje Linfático Manual en Brazos PDFDocumento3 páginasDrenaje Linfático Manual en Brazos PDFPepi Barajas AriasAún no hay calificaciones
- Anatomía Del PáncreasDocumento6 páginasAnatomía Del PáncreasD͎i͎e͎g͎o͎ A͎l͎e͎j͎a͎n͎d͎r͎o͎Aún no hay calificaciones
- Anatomia y Fisiologia de La Piel Mas TrabajodDocumento19 páginasAnatomia y Fisiologia de La Piel Mas TrabajodCristian RiquelmeAún no hay calificaciones
- Manipulaciones Drenaje Linfatico Post OperatorioDocumento3 páginasManipulaciones Drenaje Linfatico Post OperatorioLuis SanchezAún no hay calificaciones
- Enfermedades Del Sistema Linfatico 2Documento10 páginasEnfermedades Del Sistema Linfatico 2lesliculquichiconAún no hay calificaciones
- Musculos VoluntariosDocumento2 páginasMusculos VoluntariossandraAún no hay calificaciones
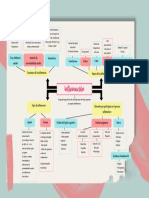
- InflamacionDocumento37 páginasInflamacionRuth ZuritaAún no hay calificaciones
- Músculos de La CaraDocumento7 páginasMúsculos de La CaraJared ZarateAún no hay calificaciones
- Tejido y Órganos LinfáticosDocumento8 páginasTejido y Órganos LinfáticosCarolina VillarroelAún no hay calificaciones
- VitekDocumento7 páginasVitekBrockamp MarggiieAún no hay calificaciones
- Los Sentidos Vista y AudiciónDocumento10 páginasLos Sentidos Vista y AudiciónYenny Liliana Rodriguez GuerreroAún no hay calificaciones
- Sistema Del Cuerpo HumanoDocumento3 páginasSistema Del Cuerpo HumanoSnader QuinterosAún no hay calificaciones
- Reflexología Zú - Vía de iniciación al Tao: Vol. IDe EverandReflexología Zú - Vía de iniciación al Tao: Vol. IAún no hay calificaciones
- Sistema LinfáticoDocumento3 páginasSistema LinfáticoAdriana PeñaAún no hay calificaciones
- Frotis Sanguneo 2Documento13 páginasFrotis Sanguneo 2María de nazaret Tovar venegasAún no hay calificaciones
- Caso 20Documento13 páginasCaso 20Coral CaroAún no hay calificaciones
- Herpes Simple y Herpes ZosterDocumento2 páginasHerpes Simple y Herpes ZosterHector Antonio Castillo SaavedraAún no hay calificaciones
- Tejido SanguineoDocumento16 páginasTejido SanguineoLeyli Tatiana Caballero VelasquezAún no hay calificaciones
- Enfermedades Metaxénicas - Salud Pública - Jaime Salazar SalazarDocumento44 páginasEnfermedades Metaxénicas - Salud Pública - Jaime Salazar Salazarluis fernandoAún no hay calificaciones
- Tarea Método BiogavalDocumento11 páginasTarea Método BiogavalnedepuAún no hay calificaciones
- Sistema Inmunitario 3esoDocumento38 páginasSistema Inmunitario 3esolauraAún no hay calificaciones
- Mapa Conceptual InflamaciónDocumento1 páginaMapa Conceptual InflamaciónMaría Fernanda Argote100% (6)
- Tema 4Documento23 páginasTema 4Pilar Burguet villenaAún no hay calificaciones
- Rol Del Laboratotio de MicobiologiaDocumento30 páginasRol Del Laboratotio de MicobiologiaConstanza FuentealbaAún no hay calificaciones
- Lupus ResumenDocumento11 páginasLupus Resumenangela arturoAún no hay calificaciones
- Tecnicas de AislamientoDocumento18 páginasTecnicas de AislamientoLuzbel DiabulusAún no hay calificaciones
- Historianaturalrinofaringitisresfriadocomn 141022193507 Conversion Gate02Documento3 páginasHistorianaturalrinofaringitisresfriadocomn 141022193507 Conversion Gate02Francisca LópezAún no hay calificaciones
- Hepatitis A Inserto)Documento2 páginasHepatitis A Inserto)Maryuri RuizAún no hay calificaciones
- N01I-2A-Párrafo Introducción y Cierre (Material) - MARZO 2020 OK (1) - 1Documento1 páginaN01I-2A-Párrafo Introducción y Cierre (Material) - MARZO 2020 OK (1) - 1Cristian JH Cosi50% (2)
- SEMANA 1, Conceptos y Definiciones de Laboratorio Clinico, SEPARATADocumento7 páginasSEMANA 1, Conceptos y Definiciones de Laboratorio Clinico, SEPARATABioRodriAún no hay calificaciones
- Practica #11 Fagocitosis y OpsonizaciónDocumento7 páginasPractica #11 Fagocitosis y OpsonizaciónAlejandro AcostaAún no hay calificaciones
- Mevacuno (3) - 1Documento2 páginasMevacuno (3) - 1Joaquin LemunAún no hay calificaciones
- Plan de ZoonosisDocumento11 páginasPlan de ZoonosisJerih Alessandro Beltran100% (2)
- TRVIH - Aspectos Técnicos y LaboratorioDocumento32 páginasTRVIH - Aspectos Técnicos y LaboratorioRicki VeAún no hay calificaciones
- Tema 10. Fiebre de Lassa. ADCDocumento7 páginasTema 10. Fiebre de Lassa. ADCkatonishiAún no hay calificaciones
- 5.traqueobronquitits Infecciosas CaninaDocumento5 páginas5.traqueobronquitits Infecciosas CaninaJonathan JeronimoAún no hay calificaciones
- Treponema Pallidum PallidumDocumento2 páginasTreponema Pallidum PallidumAngelica MartinAún no hay calificaciones
- Formato Control de Inventario LiDocumento50 páginasFormato Control de Inventario LiCentro de Salud de Subachoque EnfermeriaAún no hay calificaciones
- TBC en PediatriaDocumento23 páginasTBC en PediatriaYuriko SolisAún no hay calificaciones
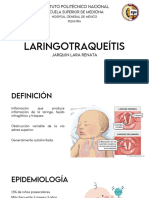
- Laringotraqueítis HGMDocumento17 páginasLaringotraqueítis HGMdra.jarquinlaraAún no hay calificaciones
- La GonorreaDocumento12 páginasLa GonorreabrendixfaAún no hay calificaciones
- Dermatosis InflamatoriasDocumento72 páginasDermatosis InflamatoriasMARIA YULIANA BERNAL DIAZAún no hay calificaciones
- CUADERNILLODocumento11 páginasCUADERNILLOJaninaAún no hay calificaciones
- ExantemasDocumento36 páginasExantemasalvaro vadell hanischAún no hay calificaciones