También podría gustarte
- CMHDocumento12 páginasCMHKarla CaballeroAún no hay calificaciones
- Capítulo 6 Sistema Del ComplementoDocumento26 páginasCapítulo 6 Sistema Del ComplementoJonatan Pinto100% (1)
- Modelos de MangasDocumento6 páginasModelos de MangasGLORIA MARITSA RAMON ASTOAún no hay calificaciones
- Guia Pinchos La LaurelDocumento80 páginasGuia Pinchos La LaurelasdfasfsAún no hay calificaciones
- Desarrollo de CuellosDocumento2 páginasDesarrollo de CuellosGLORIA MARITSA RAMON ASTOAún no hay calificaciones
- Patrón JubónDocumento18 páginasPatrón JubónMargarita100% (1)
- Astral AmDocumento16 páginasAstral AmClaudia Elena Toledo SerrallongaAún no hay calificaciones
- Modisteria Principios Basicos de Corte PDFDocumento32 páginasModisteria Principios Basicos de Corte PDFjlfigueroag6902Aún no hay calificaciones
- Alta CosturaDocumento6 páginasAlta CosturaGLORIA MARITSA RAMON ASTOAún no hay calificaciones
- Feng Shui ConsejosDocumento11 páginasFeng Shui ConsejosEstefania LaraAún no hay calificaciones
- Histología del tejido conjuntivoDocumento14 páginasHistología del tejido conjuntivoMaría José Inostroza Contreras100% (5)
- Frotis Sanguíneo - Jenny Rivera MezaDocumento9 páginasFrotis Sanguíneo - Jenny Rivera Mezaaprolab2013100% (1)
- Maquina RectaDocumento2 páginasMaquina RectaJose G Bautista TeranAún no hay calificaciones
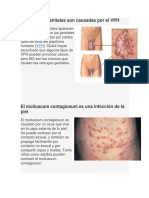
- La Verrugas Genitales Son Causadas Por El VPHDocumento2 páginasLa Verrugas Genitales Son Causadas Por El VPHCyber PlussAún no hay calificaciones
- Estructura y función del receptor de célula B y TCRDocumento9 páginasEstructura y función del receptor de célula B y TCRAnny ZhengAún no hay calificaciones
- El Material Básico para Hacer Un Maquillaje de FantasíaDocumento2 páginasEl Material Básico para Hacer Un Maquillaje de FantasíaAriana Iñiguez LuzónAún no hay calificaciones
- Analisis Microbiologicos de Pastas de Se PDFDocumento60 páginasAnalisis Microbiologicos de Pastas de Se PDFjuangabriel Jimenez SalasAún no hay calificaciones
- Tu Carta Astral Gratis, Astrologia Los ArcanosDocumento8 páginasTu Carta Astral Gratis, Astrologia Los ArcanosFernanda RiveraAún no hay calificaciones
- Miasis PDFDocumento33 páginasMiasis PDFMia Obando Velasquez100% (1)
- Práctica 2. Elaboración de Frotis SanguíneoDocumento9 páginasPráctica 2. Elaboración de Frotis SanguíneoVANESSA YARAVY MURGUIA MEDINA100% (1)
- Eosinofilos en OrinaDocumento1 páginaEosinofilos en Orinael zair100% (1)
- Autoinmunidad: respuesta inmune contra lo propioDocumento38 páginasAutoinmunidad: respuesta inmune contra lo propioVaquero Soleto Maria ReneéAún no hay calificaciones
- L1, Venopunción, Frote Sanguíneo y Tinción Giemsa y WrightDocumento15 páginasL1, Venopunción, Frote Sanguíneo y Tinción Giemsa y Wrightjackeline100% (1)
- PCR Real Tiempo Salmonella AlimentosDocumento24 páginasPCR Real Tiempo Salmonella AlimentosYessica100% (1)
- Diapositivas de EstafilococosDocumento33 páginasDiapositivas de EstafilococosMilagrOs Benites Reyes100% (1)
- ArnDocumento3 páginasArnDaniel Garcia100% (1)
- Manual de Laboratorio de MicologiaDocumento119 páginasManual de Laboratorio de MicologiaJosé Ángel Robles100% (2)
- TincionesDocumento19 páginasTincionesRafael Choque0% (1)
- Generalidades GeneticaDocumento3 páginasGeneralidades GeneticaDra Daniela TorresAún no hay calificaciones
- Seminario 5 Pruebas Cruzadas IncompatiblesDocumento16 páginasSeminario 5 Pruebas Cruzadas IncompatiblescelulasdianaAún no hay calificaciones
- Como Maquillarse Como Jack SparrowDocumento5 páginasComo Maquillarse Como Jack SparrowcomomaquillarseAún no hay calificaciones
- Exudado NasalDocumento2 páginasExudado NasalSarita MedranoAún no hay calificaciones
- 01 Historia de La MicologíaDocumento11 páginas01 Historia de La MicologíaJesus Bautisto Huichi Jara100% (1)
- Inmunología RegueyroDocumento3 páginasInmunología RegueyroKarime Fahme GonzalezAún no hay calificaciones
- Inmunodeficiencias CongénitasDocumento121 páginasInmunodeficiencias CongénitasDaniela GonzalezAún no hay calificaciones
- Proceso de UrocultivoDocumento24 páginasProceso de UrocultivoJudith Yolanda Leon OchoaAún no hay calificaciones
- INFLAMACION y FAGOCITOSISDocumento7 páginasINFLAMACION y FAGOCITOSISYoly Vasquez100% (2)
- CrioglobulinasDocumento15 páginasCrioglobulinasglodovichiAún no hay calificaciones
- Antisépticos y DesinfectantesDocumento24 páginasAntisépticos y DesinfectantesAline González RomoAún no hay calificaciones
- Aislamiento de MicroorganismosDocumento11 páginasAislamiento de Microorganismos.:("*"BLacK BuLLeT"*"):.100% (2)
- Anaerobios No EsporuladosDocumento4 páginasAnaerobios No Esporuladosangelvic73Aún no hay calificaciones
- Generalidades de MicologíaDocumento21 páginasGeneralidades de Micologíamayuchilindrina100% (4)
- CROMOBLASTOMICOSIS PaolaDocumento11 páginasCROMOBLASTOMICOSIS PaolaVanessa RoseroAún no hay calificaciones
- La sangre: funciones, componentes y grupos sanguíneosDocumento18 páginasLa sangre: funciones, componentes y grupos sanguíneosedwinAún no hay calificaciones
- S. LINFATICO CompletoDocumento13 páginasS. LINFATICO CompletoMariel Gonzalez100% (1)
- Trichosporon BeigeliiDocumento11 páginasTrichosporon BeigeliiADRIAN0% (1)
- Glosario de TerminosDocumento13 páginasGlosario de TerminosCarmen M A CavadiaAún no hay calificaciones
- Lesiones Alveolares DifusasDocumento52 páginasLesiones Alveolares DifusasCarlosAntonioMaldonadoAún no hay calificaciones
- Preguntas Formativas Patología Vasos Sanguíneos y PulmónDocumento11 páginasPreguntas Formativas Patología Vasos Sanguíneos y PulmónEveline Pulinario LorenzoAún no hay calificaciones
- Complemento Mapa ConceptualDocumento14 páginasComplemento Mapa ConceptualJDominguezBernalAún no hay calificaciones
- Neisseria, Moraxella y AcinetobacterDocumento30 páginasNeisseria, Moraxella y AcinetobacterCésar MachadoAún no hay calificaciones
- Antigenos Tumorales, Respuesta Inmunitaria Frente A Tumores e InmunoterapiaDocumento5 páginasAntigenos Tumorales, Respuesta Inmunitaria Frente A Tumores e InmunoterapiaIsrael Rene Bonilla MoreiraAún no hay calificaciones
- cUESTIONARIO bACTERIOLOGIADocumento36 páginascUESTIONARIO bACTERIOLOGIAAlex ChicaizaAún no hay calificaciones
- Chopita - Inmunidad MucosasDocumento7 páginasChopita - Inmunidad MucosasMartita JuárezAún no hay calificaciones
- Coloración PAPDocumento24 páginasColoración PAPBrenda Lia CvAún no hay calificaciones
- Parasitos Flagelados - Silvio Torres Bautsia - QBP - 602Documento48 páginasParasitos Flagelados - Silvio Torres Bautsia - QBP - 602Melquiades SebbastiamAún no hay calificaciones
- Estructura y función de las moléculas del complejo mayor de histocompatibilidad (MHCDocumento17 páginasEstructura y función de las moléculas del complejo mayor de histocompatibilidad (MHCLuis DiazzAún no hay calificaciones
- MHC Complejo Mayor HistocompatibilidadDocumento25 páginasMHC Complejo Mayor HistocompatibilidadAli Partiido100% (1)
- Complejo Mayor de Histocompatibilidad MHCDocumento9 páginasComplejo Mayor de Histocompatibilidad MHCAldo AguilarAún no hay calificaciones
- Apuntes de Inmunología - Procesamiento y Presentación de AntígenoDocumento4 páginasApuntes de Inmunología - Procesamiento y Presentación de AntígenoAlienAún no hay calificaciones
- Síndrome MetabólicoDocumento4 páginasSíndrome MetabólicoAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Yaaa Bastaaa x2Documento22 páginasYaaa Bastaaa x2Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- 6th Central Pay Commission Salary CalculatorDocumento15 páginas6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- DocumentoDocumento3 páginasDocumentoAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
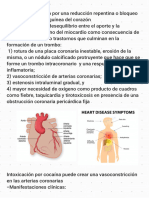
- Pericarditis Resumen HarrisonDocumento5 páginasPericarditis Resumen HarrisonAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Síndrome Coronario Agudo 230328 234219Documento8 páginasSíndrome Coronario Agudo 230328 234219Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía de Temas Anatomía 2021Documento3 páginasGuía de Temas Anatomía 2021Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía BioquiDocumento6 páginasGuía BioquiAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía de AnatoDocumento13 páginasGuía de AnatoAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía de Temas Anatomía 2021Documento3 páginasGuía de Temas Anatomía 2021Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Dermatomas ApunteDocumento1 páginaDermatomas ApunteAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Ejercicio 0.157251248Documento1 páginaEjercicio 0.157251248Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía de AnatoDocumento13 páginasGuía de AnatoAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Guía BioquiDocumento6 páginasGuía BioquiAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Manual de Laboratorio 2021Documento104 páginasManual de Laboratorio 2021Alarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Cap 1 y 8 de MooreDocumento403 páginasCap 1 y 8 de MooreAlarcon Osorio Cristhian AlfredoAún no hay calificaciones
- Libro de BiologíaDocumento864 páginasLibro de BiologíaAlarcon Osorio Cristhian Alfredo91% (35)
- Semiologia Alvarado 7 Edicion PDFDocumento531 páginasSemiologia Alvarado 7 Edicion PDFLaura Sofia89% (18)
- Examen Latin Selectividad Junio 2019 Enunciado PDFDocumento3 páginasExamen Latin Selectividad Junio 2019 Enunciado PDFsoletebcAún no hay calificaciones
- Calcular La Diferencia Entre Dos Fechas JUDITHDocumento6 páginasCalcular La Diferencia Entre Dos Fechas JUDITHjudithzapataAún no hay calificaciones
- Sesión COM Miercoles 16-11-2022 Leyenda La HuacacchinaDocumento8 páginasSesión COM Miercoles 16-11-2022 Leyenda La HuacacchinaedelmiraAún no hay calificaciones
- Taller Modulo FinancieroDocumento12 páginasTaller Modulo FinancieroádrianAún no hay calificaciones
- Perforadora de Chimenea 3Documento4 páginasPerforadora de Chimenea 3Byron Giácomo Trigoso MarínAún no hay calificaciones
- Hospital 120 CamasDocumento46 páginasHospital 120 CamasJorge SanangoAún no hay calificaciones
- Tarea 3 - Administracion ContemporáneaDocumento5 páginasTarea 3 - Administracion ContemporáneaGiovanny JairAún no hay calificaciones
- Liberacion de HogarDocumento13 páginasLiberacion de Hogarhmartinez_escobarAún no hay calificaciones
- GPC BE No 104 Manejo de Intoxicacion Aguda Por Sustancias Recreativas IGSSDocumento103 páginasGPC BE No 104 Manejo de Intoxicacion Aguda Por Sustancias Recreativas IGSSJose Fuentes100% (1)
- Excavaciones en mecánica de rocas: criterios de excavabilidad aplicados a desmontes en calizas y pizarrasDocumento34 páginasExcavaciones en mecánica de rocas: criterios de excavabilidad aplicados a desmontes en calizas y pizarrasMiguel Bedón GomezAún no hay calificaciones
- Máquina de Turing: Funcionamiento y aplicación en el código EnigmaDocumento4 páginasMáquina de Turing: Funcionamiento y aplicación en el código EnigmaDavid MoralesAún no hay calificaciones
- El Derecho Internacional AmbientalDocumento32 páginasEl Derecho Internacional AmbientalCristian VásquezAún no hay calificaciones
- Partidos Politicos 1821-1870 en GuatemalaDocumento12 páginasPartidos Politicos 1821-1870 en GuatemalaOtonielLopezAún no hay calificaciones
- Zapata - Las Sesenta y Siete Celebres Preguntas Del Teologo EspañolDocumento9 páginasZapata - Las Sesenta y Siete Celebres Preguntas Del Teologo EspañolJuan M Garcia CondeAún no hay calificaciones
- InecuacionesDocumento3 páginasInecuacionesMiraida Millán RodríguezAún no hay calificaciones
- Comparación Metodología Tradicional y Comprensiva en La Práctica Del BaloncestoDocumento16 páginasComparación Metodología Tradicional y Comprensiva en La Práctica Del Baloncestocamilo ramirezAún no hay calificaciones
- Qué Fue Primero La Gallina o El HuevoDocumento2 páginasQué Fue Primero La Gallina o El HuevoJesús GonzalezAún no hay calificaciones
- Argumentacion Juridica TP4, SIGLO 21Documento4 páginasArgumentacion Juridica TP4, SIGLO 21Ayee BravoAún no hay calificaciones
- Bacterias Ii P 2021Documento13 páginasBacterias Ii P 2021Obed GodenAún no hay calificaciones
- El DineroDocumento12 páginasEl Dineroluz angely100% (1)
- 058-Carta Cierre de Vias Feria ParqueDocumento3 páginas058-Carta Cierre de Vias Feria ParqueCarlo ManroAún no hay calificaciones
- Reseña de Elizabeth (1998)Documento4 páginasReseña de Elizabeth (1998)Jorge Luis Herrera0% (1)
- El Justo FloreceraDocumento5 páginasEl Justo FloreceraMiguel CgAún no hay calificaciones
- TAREADocumento6 páginasTAREADenisse MontoyaAún no hay calificaciones
- Unidad 3Documento6 páginasUnidad 3taniaAún no hay calificaciones
- 1.2. Es Tiempo de DecidirDocumento2 páginas1.2. Es Tiempo de DecidirDiego RhAún no hay calificaciones
- TAROT EVOLUTIVO - Primeros Pasos - Diana López IriarteDocumento12 páginasTAROT EVOLUTIVO - Primeros Pasos - Diana López IriarteAngel Jurado100% (4)
- Pasto Jaragua2Documento8 páginasPasto Jaragua2David Martin SykoraAún no hay calificaciones
- 24.2. DPM - Sentencia SincromarcasDocumento10 páginas24.2. DPM - Sentencia SincromarcasSalome Perea IsazaAún no hay calificaciones
- Ejercicio de LocalizacionDocumento9 páginasEjercicio de LocalizacionKevin Stiven Rocha MolinaAún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- ABC de los procedimientos médicos básicos: Una guía de aprendizaje y enseñanza para profesionales de medicinaDe EverandABC de los procedimientos médicos básicos: Una guía de aprendizaje y enseñanza para profesionales de medicinaCalificación: 4 de 5 estrellas4/5 (4)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- La Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoDe EverandLa Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoCalificación: 5 de 5 estrellas5/5 (2)