También podría gustarte
- Enzimas ReguladorasDocumento4 páginasEnzimas ReguladorasLucas Tapia Parra0% (1)
- Enzimas ReguladorasDocumento16 páginasEnzimas ReguladorasAurora Dannae Véliz GonzálezAún no hay calificaciones
- 5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasDocumento7 páginas5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasCortes Burgos Cielo SherlynAún no hay calificaciones
- Enzimas Reguladoras o Enzimas AlostéricasDocumento5 páginasEnzimas Reguladoras o Enzimas AlostéricasNitgma DcAún no hay calificaciones
- Enzimas AlostericasDocumento6 páginasEnzimas AlostericasEnrique IV CorderoAún no hay calificaciones
- Exposicion Bioquimica 2Documento12 páginasExposicion Bioquimica 2Jeff HubbardAún no hay calificaciones
- Regulación A Nivel EnzimáticoDocumento3 páginasRegulación A Nivel EnzimáticoMaría AhumadaAún no hay calificaciones
- Interacción AlostéricaDocumento3 páginasInteracción AlostéricaNathaly Rojas Jaramillo100% (1)
- Enzimas ReguladorasDocumento7 páginasEnzimas ReguladorasLuzMa ViñasAún no hay calificaciones
- Inhibición y Regulación EnzimáticaDocumento5 páginasInhibición y Regulación EnzimáticaLourdes SoriaAún no hay calificaciones
- Enzimas Taller de RepasoDocumento19 páginasEnzimas Taller de RepasoVerónica VelásquezAún no hay calificaciones
- Enzimas AleostericasDocumento3 páginasEnzimas AleostericasMary_Coll03Aún no hay calificaciones
- Conferencia 5.biocatalizadores. Regulación EnzDocumento45 páginasConferencia 5.biocatalizadores. Regulación EnzNoybAún no hay calificaciones
- Reguladores de La Actividad EnzimáticaDocumento6 páginasReguladores de La Actividad EnzimáticaOlenka MurielAún no hay calificaciones
- Enzimas AlostéricasDocumento3 páginasEnzimas AlostéricasJF MMAún no hay calificaciones
- Enzimas III-estrategias de Catalíticas y RegulatoriasDocumento10 páginasEnzimas III-estrategias de Catalíticas y RegulatoriasOscar eli Noreña castañoAún no hay calificaciones
- Webquest ENZIMAS Equipo CDocumento19 páginasWebquest ENZIMAS Equipo CGina RojasAún no hay calificaciones
- PREGUNTAS RESUELTAS EnzimasDocumento9 páginasPREGUNTAS RESUELTAS Enzimasstiben nova0% (1)
- Clase 4 - Regulación Actividad EnzimáticaDocumento7 páginasClase 4 - Regulación Actividad EnzimáticaanaAún no hay calificaciones
- Mecanismo de Acción de Las EnzimasDocumento7 páginasMecanismo de Acción de Las EnzimasYumekoRiverAún no hay calificaciones
- 6 Enzimas 2 AlosterismoDocumento15 páginas6 Enzimas 2 AlosterismoTony LopezAún no hay calificaciones
- E. ZimasDocumento15 páginasE. Zimasluis fernandoAún no hay calificaciones
- Enzimas Reguladoras - Dra. GilDocumento35 páginasEnzimas Reguladoras - Dra. GilHinO - Arturo100% (1)
- Enzimas III-estrategias de Catalíticas y RegulatoriasDocumento10 páginasEnzimas III-estrategias de Catalíticas y Regulatorias40. Vasquez Castro Julian DavidAún no hay calificaciones
- EnzimasDocumento4 páginasEnzimasMANUEL EDUARDO SIERRA GARCIAAún no hay calificaciones
- Marco TeoricoDocumento8 páginasMarco TeoricoSebastian AlmeidaAún no hay calificaciones
- Factores Que Afectan La Actividad EnzimáticaDocumento4 páginasFactores Que Afectan La Actividad EnzimáticaDavid DiazAún no hay calificaciones
- Respaso 1Documento17 páginasRespaso 1Aldair MedinaAún no hay calificaciones
- Bioquimica Mecanismo de RegulacionDocumento3 páginasBioquimica Mecanismo de RegulacionKarelys AbrahamAún no hay calificaciones
- La Regulación AlostéricaDocumento1 páginaLa Regulación AlostéricaMaria PalaciosAún no hay calificaciones
- Taller y Glosario de Enzimas y Factores Que Afectan Las EnzimasDocumento12 páginasTaller y Glosario de Enzimas y Factores Que Afectan Las EnzimasAngie Villarreal100% (1)
- EnzimasDocumento5 páginasEnzimasPerla FelizAún no hay calificaciones
- En ZimasDocumento4 páginasEn ZimasMaria GarciaAún no hay calificaciones
- Explicar Cuáles Son Los Principales Factores Que Afectan A La Actividad EnzimáticaDocumento7 páginasExplicar Cuáles Son Los Principales Factores Que Afectan A La Actividad EnzimáticaRuben Fonseca100% (1)
- ENZIMASDocumento28 páginasENZIMASAlfredo MéndezAún no hay calificaciones
- Regulacion EnzimaticaDocumento12 páginasRegulacion EnzimaticaKurtis JuarezAún no hay calificaciones
- Cambios AlostericosDocumento2 páginasCambios AlostericosOscar LeónAún no hay calificaciones
- Regulación AlostéricaDocumento5 páginasRegulación AlostéricaJosejuan DuranAún no hay calificaciones
- Ej Reacciones Enzimáticas y Enzimas ReguladorasDocumento2 páginasEj Reacciones Enzimáticas y Enzimas ReguladorasAdrianne FrazãoAún no hay calificaciones
- 13 AlosterismoDocumento42 páginas13 AlosterismoRuben CastilloAún no hay calificaciones
- Estructura y Mecanismos de Las Enzimas. Cofactores y Coenzimas. Función Biológica y Control de La Actividad.Documento6 páginasEstructura y Mecanismos de Las Enzimas. Cofactores y Coenzimas. Función Biológica y Control de La Actividad.Marina100% (1)
- ALOSTERICADocumento10 páginasALOSTERICAtanyAún no hay calificaciones
- Arreglado para ImprimirDocumento31 páginasArreglado para ImprimirMilton Cesar Rodrigo TantaleanAún no hay calificaciones
- Enzimologia Todo 2 ParcialDocumento90 páginasEnzimologia Todo 2 ParcialDavia BlasAún no hay calificaciones
- Catalizadores BiológicosDocumento7 páginasCatalizadores BiológicosDaniela Olea CisternasAún no hay calificaciones
- Cuestionario 4. Enzimas.Documento10 páginasCuestionario 4. Enzimas.andrea rochaAún no hay calificaciones
- Enzima BioquimicaDocumento36 páginasEnzima BioquimicaMarcos antonio suarez vergaraAún no hay calificaciones
- Cinetica EnzimáticaDocumento5 páginasCinetica EnzimáticaJANE BARAJASAún no hay calificaciones
- Información BiologiaDocumento3 páginasInformación BiologiaEmerson Tiago Camacho DíazAún no hay calificaciones
- Regulación de La Actividad Enzimatica Grupo 3Documento29 páginasRegulación de La Actividad Enzimatica Grupo 3AlexandraParedesPuertaAún no hay calificaciones
- AlosterismoDocumento12 páginasAlosterismoAbigail Becerril SandovalAún no hay calificaciones
- Para Que Una Célula Viva Se Desarrolle y ReproduzcaDocumento5 páginasPara Que Una Célula Viva Se Desarrolle y ReproduzcaElder CulajayAún no hay calificaciones
- Biologia 1121 EnzimasDocumento10 páginasBiologia 1121 Enzimasnairoby herrera smartAún no hay calificaciones
- Cinética EnzimáticaDocumento4 páginasCinética EnzimáticaDanii Castle RoblesAún no hay calificaciones
- Regulación Alostérica - José GuizarDocumento3 páginasRegulación Alostérica - José GuizarJosé Guizar AguilarAún no hay calificaciones
- GTP T1.base Molecular y Físicoquímica de La Vida 6 Parte Enzimas 2016-17Documento41 páginasGTP T1.base Molecular y Físicoquímica de La Vida 6 Parte Enzimas 2016-17Josue MantillaAún no hay calificaciones
- Practica de Catalisis (Investigacion)Documento4 páginasPractica de Catalisis (Investigacion)Jas AcostaAún no hay calificaciones
- Enzimas Reguladoras y No ReguladorasDocumento28 páginasEnzimas Reguladoras y No ReguladorasElszy MárquezAún no hay calificaciones
- DinamicaDocumento5 páginasDinamicaAdrián GpAún no hay calificaciones
- Cancer BiofarmacosDocumento1 páginaCancer BiofarmacosAdrián GpAún no hay calificaciones
- ContinueDocumento3 páginasContinueAdrián GpAún no hay calificaciones
- Reseña Dark WatersDocumento2 páginasReseña Dark WatersAdrián GpAún no hay calificaciones
- Guia Silent Hill Parte 1Documento35 páginasGuia Silent Hill Parte 1Adrián GpAún no hay calificaciones
- Cuestionario Bioogia CelularDocumento5 páginasCuestionario Bioogia CelularAdrián GpAún no hay calificaciones
- LevaduraDocumento3 páginasLevaduraAdrián GpAún no hay calificaciones
- Organizador GráficoDocumento6 páginasOrganizador GráficoAdrián GpAún no hay calificaciones
- Ejercicio QuimicaDocumento6 páginasEjercicio QuimicaAdrián GpAún no hay calificaciones
- AntiviralesDocumento3 páginasAntiviralesAdrián GpAún no hay calificaciones
- Ampliaciones de La GenéticaDocumento18 páginasAmpliaciones de La GenéticaAdrián GpAún no hay calificaciones
- Guia - Enzimas PDFDocumento8 páginasGuia - Enzimas PDFAdrián GpAún no hay calificaciones
- Turgencia y PlasmólisisDocumento5 páginasTurgencia y PlasmólisisBrayan MartínezAún no hay calificaciones
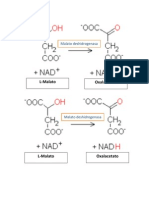
- La Enzima Malato DeshidrogenasaDocumento2 páginasLa Enzima Malato DeshidrogenasaKaren Torres JuárezAún no hay calificaciones
- 10.2 MutacionesDocumento30 páginas10.2 MutacionesCesar Eneque ChancafeAún no hay calificaciones
- Obj. 2 - TranscripciónDocumento5 páginasObj. 2 - TranscripciónCristinaAún no hay calificaciones
- Eva de Transporte CelularDocumento11 páginasEva de Transporte Celularrosana vargasAún no hay calificaciones
- Practica Calificada BioinformaticaDocumento2 páginasPractica Calificada BioinformaticaLuis Eduardo Strater Mc LellanAún no hay calificaciones
- 16 NucleoDocumento19 páginas16 NucleoGabriela Gómez CarboneAún no hay calificaciones
- Codigo GeneticoDocumento1 páginaCodigo GeneticoBrahyan SuárezAún no hay calificaciones
- Reticulo Endoplasmatico Liso y Rugoso, FuncionesDocumento5 páginasReticulo Endoplasmatico Liso y Rugoso, FuncionesAna MarcelaAún no hay calificaciones
- Regulación de La Actividad EnzimáticaDocumento6 páginasRegulación de La Actividad EnzimáticaJuan ReyesAún no hay calificaciones
- Lectura Membrana CelularDocumento1 páginaLectura Membrana CelularDiana Claros AlvarezAún no hay calificaciones
- Electroforesis de PéptidosDocumento4 páginasElectroforesis de PéptidosRAQUELAún no hay calificaciones
- Anemia Falciforme - Equipo 1Documento19 páginasAnemia Falciforme - Equipo 1Casandra Lobato DomínguezAún no hay calificaciones
- VirusDocumento9 páginasVirusJoel Max Paul Cutipa100% (1)
- Problemario FLUJO DE INFORMACIÓN GÉNICA MEFI 2022docxDocumento4 páginasProblemario FLUJO DE INFORMACIÓN GÉNICA MEFI 2022docxVero GonzalezAún no hay calificaciones
- Taller de Refuerzo 6Documento3 páginasTaller de Refuerzo 6Tatiana Ortiz50% (2)
- Citoesqueleto y Mo-Lidad Intracelular: Microtubulos y Filamentos IntermediosDocumento38 páginasCitoesqueleto y Mo-Lidad Intracelular: Microtubulos y Filamentos IntermediosAlejandro MartínezAún no hay calificaciones
- P9 TriacilgliceridosDocumento7 páginasP9 TriacilgliceridosElizabeth soto MontesAún no hay calificaciones
- Técnica de Western BlotDocumento5 páginasTécnica de Western BlotGABRIEL ALEXANDER ONCOY MOREAún no hay calificaciones
- Caracterización Molecular y Bioquímica de Una Nueva XilanasaDocumento33 páginasCaracterización Molecular y Bioquímica de Una Nueva XilanasaDiana RodríguezAún no hay calificaciones
- Historia de La Biología MolecularDocumento5 páginasHistoria de La Biología MolecularDeyaTaseAún no hay calificaciones
- TALLER No 001 GENERALIDADES ADNDocumento7 páginasTALLER No 001 GENERALIDADES ADNFreddy Santiago Navarro88% (8)
- PCR MultiplexDocumento2 páginasPCR MultiplexMatt GamerAún no hay calificaciones
- Herencia Mitocondrial PDFDocumento31 páginasHerencia Mitocondrial PDFMaria Jose Garcia BermejoAún no hay calificaciones
- Nutrientes EspecificosDocumento266 páginasNutrientes EspecificosLUCEROAún no hay calificaciones
- Informe N1 Codigo Genetico y MutacionesDocumento20 páginasInforme N1 Codigo Genetico y MutacionesJohan Joseph Romero Macavilca100% (2)
- 2 Fundamentos de Nutrición Clínica - Interrrelaciones Metabólicas 2020 PDFDocumento28 páginas2 Fundamentos de Nutrición Clínica - Interrrelaciones Metabólicas 2020 PDFcucurruscucucuAún no hay calificaciones
- Estructura Mitocondrial y FunciónDocumento5 páginasEstructura Mitocondrial y FunciónDaniela Torres ZapataAún no hay calificaciones
- Reporte de Seminario II BCyMDocumento2 páginasReporte de Seminario II BCyMGianpaolo Ricardo Lara AsencioAún no hay calificaciones
- Ensayo Del DNADocumento21 páginasEnsayo Del DNADanielLoaRaAún no hay calificaciones
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDe EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaCalificación: 5 de 5 estrellas5/5 (1875)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadDe EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadCalificación: 4.5 de 5 estrellas4.5/5 (1833)
- Recupera tu mente, reconquista tu vidaDe EverandRecupera tu mente, reconquista tu vidaCalificación: 5 de 5 estrellas5/5 (8)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (203)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)De EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Calificación: 5 de 5 estrellas5/5 (8)
- La violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaDe EverandLa violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaCalificación: 4 de 5 estrellas4/5 (2)
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaDe EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaCalificación: 4.5 de 5 estrellas4.5/5 (766)
- Resetea tu mente. Descubre de lo que eres capazDe EverandResetea tu mente. Descubre de lo que eres capazCalificación: 5 de 5 estrellas5/5 (196)
- Terapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSDe EverandTerapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (64)
- El poder del optimismo: Herramientas para vivir de forma más positivaDe EverandEl poder del optimismo: Herramientas para vivir de forma más positivaCalificación: 5 de 5 estrellas5/5 (16)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Contra la ansiedad: Una guía completa para manejar emociones difícilesDe EverandContra la ansiedad: Una guía completa para manejar emociones difícilesCalificación: 5 de 5 estrellas5/5 (58)
- DMT: La molécula del espíritu (DMT: The Spirit Molecule): Las revolucionarias investigaciones de un medico sobre la biologia de las experiencias misticas y cercanas a la muerteDe EverandDMT: La molécula del espíritu (DMT: The Spirit Molecule): Las revolucionarias investigaciones de un medico sobre la biologia de las experiencias misticas y cercanas a la muerteCalificación: 4.5 de 5 estrellas4.5/5 (19)
- Influencia. La psicología de la persuasiónDe EverandInfluencia. La psicología de la persuasiónCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Una mente en calma: Técnicas para manejar los pensamientos intrusivosDe EverandUna mente en calma: Técnicas para manejar los pensamientos intrusivosCalificación: 4.5 de 5 estrellas4.5/5 (142)
- Cómo romper el ciclo de pensamientos negativosDe EverandCómo romper el ciclo de pensamientos negativosCalificación: 4.5 de 5 estrellas4.5/5 (274)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- El Tesoro Escondido (Hidden Treasure): La vida interior de niños y adolescentes. Terapia infanto-juvenilDe EverandEl Tesoro Escondido (Hidden Treasure): La vida interior de niños y adolescentes. Terapia infanto-juvenilCalificación: 5 de 5 estrellas5/5 (4)
- No desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaDe EverandNo desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaCalificación: 5 de 5 estrellas5/5 (13)
- Ondas de choque extracorpóreas radiales: Aplicación en patologías músculo esqueléticasDe EverandOndas de choque extracorpóreas radiales: Aplicación en patologías músculo esqueléticasCalificación: 5 de 5 estrellas5/5 (3)
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaDe EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaDe EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaCalificación: 5 de 5 estrellas5/5 (2)