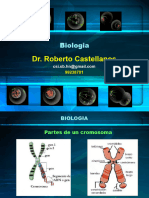
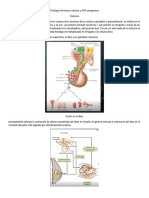
También podría gustarte
- Citologia CervicalDocumento69 páginasCitologia CervicalIsabella OjedaAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- La Teoría de Los Sistemas Aplicada A Las OrganizacionesDocumento32 páginasLa Teoría de Los Sistemas Aplicada A Las Organizacionesleidi0% (1)
- Taller EntrecruzamientoDocumento6 páginasTaller EntrecruzamientoMiriam CaroAún no hay calificaciones
- Práctica 9 Extracción de ADNDocumento4 páginasPráctica 9 Extracción de ADNAbby Valde75% (4)
- 6B. LigamientoDocumento86 páginas6B. Ligamientojoe0800% (1)
- Practica 7. Entrecruzamiento y Mapeo GeneticoDocumento13 páginasPractica 7. Entrecruzamiento y Mapeo GeneticoyamilAún no hay calificaciones
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDe EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónAún no hay calificaciones
- LigamientoDocumento90 páginasLigamientoStefy RecaldeAún no hay calificaciones
- Clase 5 Recombinacion y Genes LigadosDocumento28 páginasClase 5 Recombinacion y Genes LigadosCatalina CarrascoAún no hay calificaciones
- Genetica 5Documento16 páginasGenetica 5Anto NadinAún no hay calificaciones
- LigamientoDocumento38 páginasLigamientoapi-26818885100% (1)
- Genetica AnimalDocumento148 páginasGenetica AnimalIngrid AcevedoAún no hay calificaciones
- Genetica Ligamiento EntrecruzamientoDocumento17 páginasGenetica Ligamiento EntrecruzamientodbzoAún no hay calificaciones
- Seminario 5 GenéticaDocumento31 páginasSeminario 5 GenéticaPamela Martínez BarriosAún no hay calificaciones
- Seminario 5 EspañolDocumento33 páginasSeminario 5 EspañolJorge Chávez MegoAún no hay calificaciones
- Taller EntrecruzamientoDocumento6 páginasTaller EntrecruzamientoYARITZA VANESSA BARCELO OJEDAAún no hay calificaciones
- Psicobiología Tema 2 ResumenDocumento11 páginasPsicobiología Tema 2 ResumenArabiaDíazAún no hay calificaciones
- PRACTICA #09 Genetica VegetalDocumento10 páginasPRACTICA #09 Genetica VegetalMirian Velasquez GuarnizAún no hay calificaciones
- Tarea 9 Practica 7 LigamientoDocumento2 páginasTarea 9 Practica 7 LigamientoDennis Farroñan Espinoza100% (1)
- Tipos de Accion GenicaDocumento15 páginasTipos de Accion GenicaLeón MedinaAún no hay calificaciones
- Cuadro de PunnetDocumento47 páginasCuadro de PunnetEdwin Rodriguez DzulAún no hay calificaciones
- Johana Carolina Soto Sedano. Msc. PHD: Jcsotos@Unal - Edu.CoDocumento57 páginasJohana Carolina Soto Sedano. Msc. PHD: Jcsotos@Unal - Edu.CoDiego GarcíaAún no hay calificaciones
- Ligamiento Mapeo CromosomicoDocumento29 páginasLigamiento Mapeo CromosomicoLUIS ANTONIO GUTIERREZ BARRIALAún no hay calificaciones
- Tema 5 GmaDocumento19 páginasTema 5 GmaanabelenorcajadapastorAún no hay calificaciones
- Ligamiento Recombinación y Mapeo de Genes EucarionteDocumento5 páginasLigamiento Recombinación y Mapeo de Genes EucariontePepoHernandez100% (1)
- Genética PostmendelianaDocumento9 páginasGenética PostmendelianaVentitasSantiagoAún no hay calificaciones
- Tema 5 Ligamiento y Recombinación 2023Documento124 páginasTema 5 Ligamiento y Recombinación 2023mrr00133Aún no hay calificaciones
- Ligamiento y TiposDocumento2 páginasLigamiento y TiposRut Juliette PacherresAún no hay calificaciones
- GuiaProblema1ercuat2014 1aparteDocumento26 páginasGuiaProblema1ercuat2014 1apartePaulie MuñozAún no hay calificaciones
- Ligamientoentrecruzamiento 150126175747 Conversion Gate01Documento12 páginasLigamientoentrecruzamiento 150126175747 Conversion Gate01Diego LopezAún no hay calificaciones
- PolihibridoDocumento30 páginasPolihibridoCharlin Cerna RuizAún no hay calificaciones
- GENÉTICADocumento18 páginasGENÉTICAGustavo HernándezAún no hay calificaciones
- Ejercicios de LigamientoDocumento14 páginasEjercicios de LigamientoPaula S VargasAún no hay calificaciones
- Variabilidad Genética y Recombinación.Documento8 páginasVariabilidad Genética y Recombinación.Carolina Rivera ReyAún no hay calificaciones
- Seminario 5 CompletoDocumento11 páginasSeminario 5 CompletoLeslie Gonzales ChipanaAún no hay calificaciones
- BiologiaDocumento20 páginasBiologiaRoberto CastellanosAún no hay calificaciones
- 09 Ligamiento Mapeo CromosomicoDocumento43 páginas09 Ligamiento Mapeo CromosomicoJawy RamírezAún no hay calificaciones
- 17-Ligamiento y Recombinación en EucariontesDocumento25 páginas17-Ligamiento y Recombinación en EucariontesAlmudena MeiAún no hay calificaciones
- Resumen Segundo ParcialDocumento13 páginasResumen Segundo ParcialRocio DabosAún no hay calificaciones
- 2-Tipos de Accion GenicaDocumento15 páginas2-Tipos de Accion GenicaNoelia Alejandra Begazo AbrilAún no hay calificaciones
- Bases Genéticas para El Análisis de Ligamento y AsociaciónDocumento9 páginasBases Genéticas para El Análisis de Ligamento y Asociaciónmartin francoAún no hay calificaciones
- Genetic ADocumento4 páginasGenetic AGertrudis Martinez VazquezAún no hay calificaciones
- GeneticaDocumento6 páginasGeneticaNicolas SierraAún no hay calificaciones
- Ligamentos GeneticosDocumento3 páginasLigamentos GeneticosdianaAún no hay calificaciones
- Genética - Diego MeléndezDocumento29 páginasGenética - Diego Meléndezdiego melendezAún no hay calificaciones
- Practica 6Documento3 páginasPractica 6Jonathan RaxcacóAún no hay calificaciones
- HERENCIADocumento4 páginasHERENCIAleonorAún no hay calificaciones
- Genética Mendeliana 1Documento44 páginasGenética Mendeliana 1Germain Rider FigueroaAún no hay calificaciones
- Taller 0 Repaso Biología CelularDocumento6 páginasTaller 0 Repaso Biología Celularmayelly villotaAún no hay calificaciones
- Practica #10Documento16 páginasPractica #10DINA VALERIA CAMPOSANO BARRIENTOSAún no hay calificaciones
- Mapas Genéticos y FísicosDocumento7 páginasMapas Genéticos y FísicosmatateteAún no hay calificaciones
- Codominancia. Series Alélicas. Ligamiento y RecombinaciónDocumento41 páginasCodominancia. Series Alélicas. Ligamiento y RecombinaciónFrancisco GomezAún no hay calificaciones
- Mapeo y Ligamiento - II.23Documento26 páginasMapeo y Ligamiento - II.23victorcondor666Aún no hay calificaciones
- Clase 4 de GeneticaDocumento25 páginasClase 4 de GeneticacharlisvelayarceAún no hay calificaciones
- Introducción A La Genética Médica. 13. Ligamiento y RecombinaciónDocumento25 páginasIntroducción A La Genética Médica. 13. Ligamiento y Recombinaciónmes_reis100% (1)
- Genética No MendelianaDocumento28 páginasGenética No MendelianaAlexander Oviedo BenitezAún no hay calificaciones
- Guía de Estudio Mecanismos de La Herencia (Parte II) 2021Documento8 páginasGuía de Estudio Mecanismos de La Herencia (Parte II) 2021Miyer RiveroAún no hay calificaciones
- Taller BiologiaDocumento20 páginasTaller BiologiaCristhian Andres Rojas GrisalesAún no hay calificaciones
- Herencia No MendelianaDocumento4 páginasHerencia No MendelianaEdwin VianaAún no hay calificaciones
- Mendel 1Documento17 páginasMendel 1Denisse SepúlvedaAún no hay calificaciones
- Taller 1 y 2 Biologia Celular y MolecularDocumento53 páginasTaller 1 y 2 Biologia Celular y MolecularVanessa NúñezAún no hay calificaciones
- Naturaleza Del Gen y El GenomaDocumento31 páginasNaturaleza Del Gen y El GenomaisabelAún no hay calificaciones
- Ascomicetos Division AscomycotaDocumento41 páginasAscomicetos Division AscomycotaFátima Arias Cabrejos50% (2)
- Metabolismo Del AzufreDocumento16 páginasMetabolismo Del AzufreGianella Lucely Rodriguez Andrade0% (1)
- Guia #7 Biologia 70s PDFDocumento3 páginasGuia #7 Biologia 70s PDFDIANA PATRICIA ROCHA-ACEVEDOAún no hay calificaciones
- Botánica-Tema 1Documento12 páginasBotánica-Tema 1Mariela GuevaraAún no hay calificaciones
- 3 Resumen GastrulacionDocumento10 páginas3 Resumen GastrulacionPablo AbolskyAún no hay calificaciones
- Biologia de Miller y Levien 3Documento34 páginasBiologia de Miller y Levien 3andres acevedoAún no hay calificaciones
- Alfred WallaceDocumento1 páginaAlfred WallaceYpAún no hay calificaciones
- Problemas Practica 3Documento2 páginasProblemas Practica 3brayan gutierrezAún no hay calificaciones
- Diagnóstico de Una Enfermedad GenéticaDocumento4 páginasDiagnóstico de Una Enfermedad GenéticaAdriana ProañoAún no hay calificaciones
- Digestivo IIDocumento49 páginasDigestivo IIFranciscoFiscarelliAún no hay calificaciones
- Pa3 - 2020-1 CGQC ZuDocumento12 páginasPa3 - 2020-1 CGQC Zukoguis zupaAún no hay calificaciones
- Hongos FilamentososDocumento3 páginasHongos FilamentososErendiiraValenzuelaAún no hay calificaciones
- Fisiología Hormonas Oxitocina y ADHDocumento4 páginasFisiología Hormonas Oxitocina y ADHOscar Daniel RomeroAún no hay calificaciones
- Resumen. Clasificación de Seres VivosDocumento7 páginasResumen. Clasificación de Seres VivosValerieAún no hay calificaciones
- Ecologia Del ParasitismoDocumento17 páginasEcologia Del ParasitismoArmando RodriguezAún no hay calificaciones
- Criterios de Inicio de TargaDocumento35 páginasCriterios de Inicio de TargadocalbertoAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Unidad 3 - Tarea 4 - Crecimiento MicrobianoDocumento8 páginasGuía de Actividades y Rúbrica de Evaluación - Unidad 3 - Tarea 4 - Crecimiento Microbianoeduardo arenasAún no hay calificaciones
- Guia CuartoDocumento41 páginasGuia CuartoJOSE DOBER LUCUMI URRUTIAAún no hay calificaciones
- Capitulo 5 BiodversidadDocumento28 páginasCapitulo 5 BiodversidaddavidAún no hay calificaciones
- Laboratorio MorfofisiologiaDocumento17 páginasLaboratorio MorfofisiologiaJAWERAún no hay calificaciones
- CIENCIAS BIOLÓGICAS 4TO AÑO Tema 7 ListoDocumento31 páginasCIENCIAS BIOLÓGICAS 4TO AÑO Tema 7 ListoBryanAún no hay calificaciones
- Botanic ADocumento15 páginasBotanic ACarol HernandezAún no hay calificaciones
- Reino Plantae - FERNANDODocumento5 páginasReino Plantae - FERNANDOChristian Fernández ÁlvarezAún no hay calificaciones
- Prueba Final Primero Medio 2017Documento12 páginasPrueba Final Primero Medio 2017Mauricio Javier Cuadros QuintanaAún no hay calificaciones
- Tarea 5.1 Tejido Sanguíneo HISTO 1 DR LUIS PEREZ MENDEZDocumento3 páginasTarea 5.1 Tejido Sanguíneo HISTO 1 DR LUIS PEREZ MENDEZDuveily AcostaAún no hay calificaciones