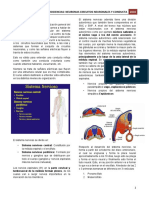
También podría gustarte
- Finaaaal Informe Bioquimica 4Documento11 páginasFinaaaal Informe Bioquimica 4Joaquin Araya MoubarakAún no hay calificaciones
- Briceño, S. Villegas, C.Documento9 páginasBriceño, S. Villegas, C.Catalina Alejandra VillegasAún no hay calificaciones
- Curva de Calibración BSA y ONFDocumento9 páginasCurva de Calibración BSA y ONFbarbaraAún no hay calificaciones
- Técnica Patch ClampDocumento3 páginasTécnica Patch ClampAlejandroHidalgoNavaAún no hay calificaciones
- Conducción Del Impulso Nervioso Por El AxónDocumento16 páginasConducción Del Impulso Nervioso Por El AxónVale LeroyerAún no hay calificaciones
- Medició de Parametros Termodinamicos PilaDocumento9 páginasMedició de Parametros Termodinamicos PilaDiego Ignacio Valderrama Soto100% (1)
- Resumen Neurobiología FINALDocumento98 páginasResumen Neurobiología FINALCynthia ScottiAún no hay calificaciones
- PH PracticaDocumento9 páginasPH PracticaeuclidesraulAún no hay calificaciones
- Reporte Laboratorio 1Documento9 páginasReporte Laboratorio 1Cristopher Lima BucioAún no hay calificaciones
- Estudio Biofisico de Los Canales IonicosDocumento8 páginasEstudio Biofisico de Los Canales IonicospedroAún no hay calificaciones
- Reporte Potencial de Membrana en Reposo PDFDocumento4 páginasReporte Potencial de Membrana en Reposo PDFAndreaNathalieChavez100% (1)
- Laboratorio 1Documento10 páginasLaboratorio 1JF MM100% (1)
- Fuerza Iónica y CoeficienteDocumento12 páginasFuerza Iónica y CoeficienteMaria Angelica Oviedo TorresAún no hay calificaciones
- Practica Compuestos de CoordinacionDocumento2 páginasPractica Compuestos de CoordinacionEva Mariana Torres0% (1)
- Qoi Unab 01Documento41 páginasQoi Unab 01Necroz182Aún no hay calificaciones
- Exercices Resolves Del AtkinsDocumento5 páginasExercices Resolves Del AtkinsRafael Ricardo Celin ManceraAún no hay calificaciones
- Guía para Usar El Simulador HHSIM PDFDocumento4 páginasGuía para Usar El Simulador HHSIM PDFJose Manuel Limia PérezAún no hay calificaciones
- Ayudant A N 4 Ligamiento en EucariontesDocumento59 páginasAyudant A N 4 Ligamiento en EucariontesBest gamer lasAún no hay calificaciones
- Primer Parcial IBMC-2015Documento4 páginasPrimer Parcial IBMC-2015LaraBen79100% (1)
- Neurociencias Clase I Neuronas Circuitos Neuronales y ConductaDocumento19 páginasNeurociencias Clase I Neuronas Circuitos Neuronales y ConductaMacarena CepedaAún no hay calificaciones
- Pract 4Documento7 páginasPract 4Fernanda PalestinaAún no hay calificaciones
- Práctica de Biología Básica. Relación Área VolumenDocumento5 páginasPráctica de Biología Básica. Relación Área Volumendiana0morales_2Aún no hay calificaciones
- Informe EquilibrioDocumento13 páginasInforme EquilibrioEstefany Duran LondoñoAún no hay calificaciones
- Potencial de Difusion y Potencial de LesionDocumento13 páginasPotencial de Difusion y Potencial de LesionLatika CzAún no hay calificaciones
- Determinacion de Aminoacidos TerminalesDocumento2 páginasDeterminacion de Aminoacidos TerminalesJeeniffer Cookiee100% (1)
- Guía de Trabajo ClonaciónDocumento6 páginasGuía de Trabajo ClonaciónMaria Catalina Guzman GelvezAún no hay calificaciones
- Practica 4Documento8 páginasPractica 4angie creyesAún no hay calificaciones
- Practica1 BioqDocumento2 páginasPractica1 BioqKari PaysAún no hay calificaciones
- 1er Parcial IBMC 2015 - Respuestas EsperadasDocumento3 páginas1er Parcial IBMC 2015 - Respuestas EsperadasLaraBen79Aún no hay calificaciones
- Ventajas y limitaciones de la acilación de Friedel-CraftsDocumento55 páginasVentajas y limitaciones de la acilación de Friedel-CraftsDARIOAún no hay calificaciones
- Clase Practica 18Documento7 páginasClase Practica 18Kenya SantacruzAún no hay calificaciones
- QuimicaDocumento8 páginasQuimicaBrën CooperAún no hay calificaciones
- Práctica 5 FisioDocumento11 páginasPráctica 5 FisioMoises Tapia BaezAún no hay calificaciones
- Fenómeno de Coactivación Muscular Durante Contracciones Isométricas Máximas PDFDocumento4 páginasFenómeno de Coactivación Muscular Durante Contracciones Isométricas Máximas PDFEzequiel LibradoAún no hay calificaciones
- GPCRs, NO, CALCIO Y PROSTAGLANDINASDocumento3 páginasGPCRs, NO, CALCIO Y PROSTAGLANDINASYoali Asiain LeconaAún no hay calificaciones
- Seminario Biopotenciales FisiologiaDocumento9 páginasSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- Laboratorio de Biología Celular - Práctica No. 8Documento4 páginasLaboratorio de Biología Celular - Práctica No. 8AndresGarcíaTorresAún no hay calificaciones
- Condensacion Aldolica Aldehidos y CetonasDocumento4 páginasCondensacion Aldolica Aldehidos y CetonasLuis MoralesAún no hay calificaciones
- Guía de Trabajo Clonación II 202120Documento5 páginasGuía de Trabajo Clonación II 202120Maria Catalina Guzman GelvezAún no hay calificaciones
- A1-Acido y BaseDocumento17 páginasA1-Acido y BaseKOBRINJAAún no hay calificaciones
- Reacciones de Proteínas IDocumento3 páginasReacciones de Proteínas IEmriel96100% (1)
- Cues Ti Ones de BioquímicaDocumento2 páginasCues Ti Ones de Bioquímicaapi-26645842Aún no hay calificaciones
- Glicólisis y FermentaciónDocumento67 páginasGlicólisis y FermentaciónDiego German R Aguilar0% (2)
- Actividad de La Membrana CelularDocumento8 páginasActividad de La Membrana CelularDiana de la CruzAún no hay calificaciones
- BiopotencialDocumento56 páginasBiopotencialMargarita Lujan AyalaAún no hay calificaciones
- Membrana PlasmaticaDocumento14 páginasMembrana PlasmaticaJosefina Cortes100% (2)
- PRACTICA 4 Potencial de Lesion y DifusionDocumento11 páginasPRACTICA 4 Potencial de Lesion y DifusionGibran Meza CastrejonAún no hay calificaciones
- Técnicas analíticas de separación. Electroforesis CapilarDocumento2 páginasTécnicas analíticas de separación. Electroforesis CapilarDani MateoAún no hay calificaciones
- PRÁCTICA No 5Documento6 páginasPRÁCTICA No 5Ale Garduño100% (1)
- Practica 15Documento7 páginasPractica 15Isaac VeraAún no hay calificaciones
- Reacciones de Edman determinan secuencia proteínasDocumento6 páginasReacciones de Edman determinan secuencia proteínasJorge Andres Espinoza100% (1)
- Tiselius y La ElectroforesisDocumento26 páginasTiselius y La ElectroforesismireAún no hay calificaciones
- Metabolismo de CarboidratosDocumento23 páginasMetabolismo de CarboidratosVerónica VelásquezAún no hay calificaciones
- Practica 1. SangerDocumento4 páginasPractica 1. SangerApolinar Aguilar BautistaAún no hay calificaciones
- Bioquímica Parciales 1 A 4Documento18 páginasBioquímica Parciales 1 A 4Frida Elisa Flores MoraAún no hay calificaciones
- Análisis de La Técnica Práctica 10.. 9Documento4 páginasAnálisis de La Técnica Práctica 10.. 9Edith AlmanzaAún no hay calificaciones
- Permeabilidad de La Membrana CelularDocumento5 páginasPermeabilidad de La Membrana CelularCamila ContrerasAún no hay calificaciones
- Practica 4 Efecto FotoelectricoDocumento5 páginasPractica 4 Efecto FotoelectricoCesar CastillogonzalezAún no hay calificaciones
- Resumen Proteínas.Documento34 páginasResumen Proteínas.Alejandra Sofía CamañoAún no hay calificaciones
- Informe 1 Lab BioquimicaDocumento10 páginasInforme 1 Lab BioquimicaCristóbal Ignacio WittersheimAún no hay calificaciones
- Diapositivas Unidad LLDocumento17 páginasDiapositivas Unidad LLivan Rodriguez RodriguezAún no hay calificaciones
- Ejercicio Codigo GeneticoDocumento3 páginasEjercicio Codigo GeneticoValentinaAún no hay calificaciones
- Wuolah Free Tema 14 BioquimicaDocumento4 páginasWuolah Free Tema 14 BioquimicaPacoSanchezBorregoAún no hay calificaciones
- Activación de los aminoácidos durante la traducciónDocumento7 páginasActivación de los aminoácidos durante la traducciónJ Mora GañanAún no hay calificaciones
- TAREA en CLASES Identifica Los Procesos Que Describe La Imagen BASES NITROGENADASDocumento5 páginasTAREA en CLASES Identifica Los Procesos Que Describe La Imagen BASES NITROGENADASYari ZambranoAún no hay calificaciones
- Aminoácidos, Péptidos y ProteínasDocumento79 páginasAminoácidos, Péptidos y Proteínasmario calaguaAún no hay calificaciones
- Ensayo ExperimentoDocumento3 páginasEnsayo ExperimentoValerie Cordoba TeranAún no hay calificaciones
- Test Enzimas Ácidos Nucl MCM 2 Bach Oct 2006Documento1 páginaTest Enzimas Ácidos Nucl MCM 2 Bach Oct 2006Prof. Víctor M. VitoriaAún no hay calificaciones
- Guia de BioquimicaDocumento11 páginasGuia de Bioquimicayaimara gonzalez leonAún no hay calificaciones
- Proteinas 2024Documento23 páginasProteinas 2024Fiorela Leonor Quiroz TurpoAún no hay calificaciones
- Foto Hidrolisis de SueroDocumento137 páginasFoto Hidrolisis de Suerojesus lopezAún no hay calificaciones
- Tema 5. EnzimologiaDocumento40 páginasTema 5. EnzimologiaMauricio Hidalgo O. PhDAún no hay calificaciones
- Mecanismo de Acción de Hormonal y Segundos Mensajeros.Documento2 páginasMecanismo de Acción de Hormonal y Segundos Mensajeros.Pablo Vega MAún no hay calificaciones
- MANUAL LAB Biología Pagina 67,68 y 69Documento3 páginasMANUAL LAB Biología Pagina 67,68 y 69Carmen De la RosaAún no hay calificaciones
- 50 Cosas Sobre EnzimasDocumento2 páginas50 Cosas Sobre EnzimasLuis EscobarAún no hay calificaciones
- Bioquimica - ApuntesDocumento181 páginasBioquimica - ApuntesPedro Henrique Gonzaga de AlmeidaAún no hay calificaciones
- Evolucion y Biomoleculas UNIDAD 1 TAREA 2Documento13 páginasEvolucion y Biomoleculas UNIDAD 1 TAREA 2ANGELA PATRICIA OCORO CRISTOBALAún no hay calificaciones
- Mapa Mental #8 - Regulación Enzimática - UasdDocumento2 páginasMapa Mental #8 - Regulación Enzimática - UasdFulanito de TalAún no hay calificaciones
- Ga 4Documento2 páginasGa 4JOSUE EDMUNDO BENDEZU AGUILARAún no hay calificaciones
- Proteinas 4 MedioDocumento39 páginasProteinas 4 MedioSumikoAún no hay calificaciones
- Guia de Trabajo ARN Importancia Biológica Del ARN, Estructura y FunciónDocumento3 páginasGuia de Trabajo ARN Importancia Biológica Del ARN, Estructura y FunciónJGP GAMERAún no hay calificaciones
- Replicación Del ADNDocumento55 páginasReplicación Del ADNDanteGomezAún no hay calificaciones
- Aminoácidos y ProteinasDocumento6 páginasAminoácidos y ProteinasFernanda RoseroAún no hay calificaciones
- Aminoácidos y estructura proteínasDocumento16 páginasAminoácidos y estructura proteínasSergio Cuevas TellecheaAún no hay calificaciones
- Gimnasio de los Cerros: Replicación, transcripción y traducción del código genéticoDocumento2 páginasGimnasio de los Cerros: Replicación, transcripción y traducción del código genéticosantiago ARAún no hay calificaciones
- BioinformáticaDocumento9 páginasBioinformáticaCarolinaAún no hay calificaciones
- Evaluación final de Biología sobre procesos genéticosDocumento5 páginasEvaluación final de Biología sobre procesos genéticosCARLOS VARGASAún no hay calificaciones
- Priones: Proteínas infecciosas con múltiples conformacionesDocumento39 páginasPriones: Proteínas infecciosas con múltiples conformacionesFaviolaValdezAún no hay calificaciones
- Análisis lácteos prácticosDocumento144 páginasAnálisis lácteos prácticosWilo MixAún no hay calificaciones
- Ficha Técnica Enterex Hepatic 90000Documento2 páginasFicha Técnica Enterex Hepatic 90000Jose MoraAún no hay calificaciones