También podría gustarte
- Anatomía FoliarDocumento26 páginasAnatomía FoliarGui LleAún no hay calificaciones
- Anatomía de La HojaDocumento71 páginasAnatomía de La HojaAna GutiérrezAún no hay calificaciones
- Anatomía de La HojaDocumento20 páginasAnatomía de La HojaLuciana Sossiré Vásquez Regalado100% (1)
- Hojas de DicotiledóneasDocumento10 páginasHojas de DicotiledóneasRocsana QGAún no hay calificaciones
- Hoja InternaDocumento3 páginasHoja Internamiguel angel tejerinaAún no hay calificaciones
- Células de Plantas y Organización VegetalDocumento9 páginasCélulas de Plantas y Organización VegetalA & C PRODUCTIONSAún no hay calificaciones
- Estructura Anatómica de La HojaDocumento13 páginasEstructura Anatómica de La Hojakovac lucasAún no hay calificaciones
- Estructura Del TalloDocumento14 páginasEstructura Del TalloDiana CBAún no hay calificaciones
- Guía N°5:: Tejidos Primarios en Hojas de Plantas VascularesDocumento4 páginasGuía N°5:: Tejidos Primarios en Hojas de Plantas VascularesVictor CofréAún no hay calificaciones
- Características de Plantas AcuáticasDocumento6 páginasCaracterísticas de Plantas AcuáticasBrendita RamirezAún no hay calificaciones
- Diagrama de Tallo y Hoja Grupo 5 (Anatomia Vegetal)Documento2 páginasDiagrama de Tallo y Hoja Grupo 5 (Anatomia Vegetal)Mary CastilloAún no hay calificaciones
- Estructura de La Hoja PDFDocumento30 páginasEstructura de La Hoja PDFromig81Aún no hay calificaciones
- Anatomia - Informe 6Documento31 páginasAnatomia - Informe 6ivanmoran938Aún no hay calificaciones
- Ob Tener Arch Ivo Recur So 2Documento41 páginasOb Tener Arch Ivo Recur So 2Cindy Leslie Pereyra NogueraAún no hay calificaciones
- Tejido EpidérmicoDocumento7 páginasTejido EpidérmicoFermin Rafael GalindoAún no hay calificaciones
- Texto 5 HOJADocumento8 páginasTexto 5 HOJAAnonymous laFvVrAún no hay calificaciones
- OrganografiaDocumento20 páginasOrganografiaElizabeth SalvadorAún no hay calificaciones
- La Hoja - BiologiaDocumento60 páginasLa Hoja - BiologiaJayJoseAndradeAún no hay calificaciones
- La HojaDocumento9 páginasLa HojaSylvia EspinAún no hay calificaciones
- La RaízDocumento6 páginasLa RaízRebeca MedinaAún no hay calificaciones
- Resumen HojaDocumento6 páginasResumen HojaemiliaAún no hay calificaciones
- El Tallo...Documento10 páginasEl Tallo...lorenandAún no hay calificaciones
- HojaDocumento12 páginasHojaZoeAún no hay calificaciones
- Tejido DérmicoDocumento8 páginasTejido Dérmicojesusabelarce50% (4)
- Aparato Lenticelas Del ArbolDocumento19 páginasAparato Lenticelas Del Arbolchampirasta100% (1)
- Morfologia Interna de RaizDocumento25 páginasMorfologia Interna de Raizm4rcucholnm50% (2)
- Parénquima VegetalDocumento5 páginasParénquima VegetalElias Alberto Flores CallesAún no hay calificaciones
- Organización Interna Del Tallo 1Documento4 páginasOrganización Interna Del Tallo 1Gustavo TulaAún no hay calificaciones
- Clases Botanica C-TalloDocumento45 páginasClases Botanica C-TalloJulio Roberto Sánchez SamayoaAún no hay calificaciones
- PALMERADocumento13 páginasPALMERAkarla Andrea Araya CruzAún no hay calificaciones
- Morfología y Anatomía de La RaízDocumento5 páginasMorfología y Anatomía de La Raízdaniela florez100% (1)
- Anatomia de La HojaDocumento3 páginasAnatomia de La HojaRebequitaLinaresAún no hay calificaciones
- QWQWQDocumento37 páginasQWQWQAbner Acuña CarrascoAún no hay calificaciones
- Estructura Primaria de RaizDocumento24 páginasEstructura Primaria de RaizEmi SerranoAún no hay calificaciones
- Oganos VegetalesDocumento44 páginasOganos VegetalesOlga MalaverAún no hay calificaciones
- Tejido EpidermicoDocumento9 páginasTejido EpidermicoLuciana Sossiré Vásquez RegaladoAún no hay calificaciones
- Botanica 3 InfoDocumento7 páginasBotanica 3 InfoFRANK CLEVER ARIMANA BARRANTESAún no hay calificaciones
- PalmeraDocumento12 páginasPalmeraRolando LinoAún no hay calificaciones
- PRACTICA 5.hoja - Tallo - RaizDocumento7 páginasPRACTICA 5.hoja - Tallo - RaizLuisa HualpaAún no hay calificaciones
- 6 Raz Morfo y Estruct Primaria PDFDocumento7 páginas6 Raz Morfo y Estruct Primaria PDFcarls vaca100% (1)
- Diapositivas Anatomia y Morfologia de Las Plantas VascularesDocumento56 páginasDiapositivas Anatomia y Morfologia de Las Plantas VascularesKary Orozco50% (2)
- Anatomía Vegetal 2da ParteDocumento38 páginasAnatomía Vegetal 2da ParteTomasAún no hay calificaciones
- Biologia Vegetal-Anatomía de La Hoja-2011Documento35 páginasBiologia Vegetal-Anatomía de La Hoja-2011maribeacaribensisAún no hay calificaciones
- Unidad 4 BotanicaDocumento12 páginasUnidad 4 BotanicaNubia HernandezAún no hay calificaciones
- Anatomía de RaízDocumento135 páginasAnatomía de RaízAndré Gamboa0% (1)
- Nuevodocumentaldelashojas 100106143624 Phpapp01Documento10 páginasNuevodocumentaldelashojas 100106143624 Phpapp01Iván AlbertoAún no hay calificaciones
- Hoja de PalmaDocumento6 páginasHoja de Palmafami retoAún no hay calificaciones
- Glosario de Términos.Documento7 páginasGlosario de Términos.Fernando TeneAún no hay calificaciones
- 3 Clase 3 Parenquima, Esclerenquima y ColenquimaDocumento33 páginas3 Clase 3 Parenquima, Esclerenquima y ColenquimaKathy Alvarez HuaytanAún no hay calificaciones
- Botanica HojasDocumento8 páginasBotanica HojasdbzoAún no hay calificaciones
- Unidad 2Documento13 páginasUnidad 2Caterina DureAún no hay calificaciones
- Las Plantas Version 2021Documento21 páginasLas Plantas Version 2021Lucia CabezonAún no hay calificaciones
- Morfologia de PlantasDocumento43 páginasMorfologia de PlantasAndres Manuel Villalobos CastroAún no hay calificaciones
- Anatomía de RaízDocumento27 páginasAnatomía de RaízAlejandro BlasAún no hay calificaciones
- Anatomía de Raíz PDFDocumento27 páginasAnatomía de Raíz PDFMarvin R. GómezAún no hay calificaciones
- El TalloDocumento16 páginasEl TalloMishaly PalaciosAún no hay calificaciones
- Cuadro Raiz y TalloDocumento10 páginasCuadro Raiz y TalloAdriana Linares100% (5)
- Exposcion Botanica TalloDocumento34 páginasExposcion Botanica TalloELIEL GONZALEZ SOLEDADAún no hay calificaciones
- Brogger 2010Documento163 páginasBrogger 2010Juan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
- Carlos Paredes Franz Cardoso y Juan TarazonaDocumento9 páginasCarlos Paredes Franz Cardoso y Juan TarazonaJuan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
- Clark, H.L. (1910) - The Echinoderms of PeruDocumento7 páginasClark, H.L. (1910) - The Echinoderms of PeruJuan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
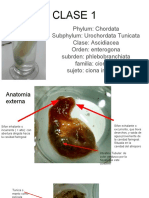
- CLASE 1 UrocordadosDocumento29 páginasCLASE 1 UrocordadosJuan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
- CLASE 1. PROT. y VERT, ANAM. - UNMSM - Hemichordata. 2017-I PDFDocumento238 páginasCLASE 1. PROT. y VERT, ANAM. - UNMSM - Hemichordata. 2017-I PDFJuan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
- Guerrero Quitorán Juan Manuel Jesús Práctica N°01 Asi Se Hizo La Tierra Animación: El Clima de La TierraDocumento11 páginasGuerrero Quitorán Juan Manuel Jesús Práctica N°01 Asi Se Hizo La Tierra Animación: El Clima de La TierraJuan Manuel Jesús Guerrero QuitoránAún no hay calificaciones
- Cultivo de Cactus y SuculentasDocumento77 páginasCultivo de Cactus y SuculentasVania Carolina91% (22)
- Viveros ForestalesDocumento14 páginasViveros ForestalesVerónika Huaire DamiánAún no hay calificaciones
- Evapotranspiracion Teoria y Calculo de ETP Por Diversos Metodos Thornthwaite Pennman Blanney Criddle HargreavesDocumento46 páginasEvapotranspiracion Teoria y Calculo de ETP Por Diversos Metodos Thornthwaite Pennman Blanney Criddle HargreavesGüido HuariAún no hay calificaciones
- Modulo CT Biología II 6Documento2 páginasModulo CT Biología II 6Rosario Yomara Rios TrujilloAún no hay calificaciones
- Informe 1 Botanica MeristematicoDocumento5 páginasInforme 1 Botanica MeristematicoArturoAún no hay calificaciones
- Bot - Práctica 1 - CASTRO - DURAN, Jean - PieerDocumento3 páginasBot - Práctica 1 - CASTRO - DURAN, Jean - PieerPieers Castro DuranAún no hay calificaciones
- Adaptaciones de Las Plantas Xerófitas A La SequíaDocumento9 páginasAdaptaciones de Las Plantas Xerófitas A La SequíaJean PajueloAún no hay calificaciones
- 00019626Documento66 páginas00019626nachinrubioAún no hay calificaciones
- 9 Riego Profesores PDFDocumento52 páginas9 Riego Profesores PDFfrcarras11Aún no hay calificaciones
- Botanica PracticaDocumento8 páginasBotanica PracticaVidal BarbozaAún no hay calificaciones
- Tipos de EstomasDocumento2 páginasTipos de EstomasAilleLiclaAún no hay calificaciones
- ArtículoDocumento3 páginasArtículojaugustonmAún no hay calificaciones
- Evaluacion 1 Nutri VegDocumento32 páginasEvaluacion 1 Nutri VegrugopaAún no hay calificaciones
- Mecanismos Fisiologicos de Apertura y Cierre de Los EstomasDocumento17 páginasMecanismos Fisiologicos de Apertura y Cierre de Los EstomasPepe Gonzales Escobar0% (1)
- Estrés Por Sequia en PlantasDocumento4 páginasEstrés Por Sequia en PlantasAme Mena SantosAún no hay calificaciones
- Bien Este para FloresDocumento127 páginasBien Este para FloresPepe Gonzales EscobarAún no hay calificaciones
- Caracteres Epidermicos de Hojas de Singonio PDFDocumento8 páginasCaracteres Epidermicos de Hojas de Singonio PDFHugomovichAún no hay calificaciones
- Rp-Cta4 - k08 - FichaDocumento10 páginasRp-Cta4 - k08 - Ficharosa luz100% (2)
- Laboratorio 4Documento9 páginasLaboratorio 4SebastianBojacaAún no hay calificaciones
- 002 Prácticas LaboratorioDocumento4 páginas002 Prácticas Laboratoriotania bolañosAún no hay calificaciones
- PRACTICA 4 Estomas y Medida de La TranspiracionDocumento12 páginasPRACTICA 4 Estomas y Medida de La TranspiracionJosé LuisAún no hay calificaciones
- GUIA FOTOSINTESIS Resumen de ContenidoDocumento8 páginasGUIA FOTOSINTESIS Resumen de ContenidoCarolina Troncoso RetamalAún no hay calificaciones
- Capsicum AnnumDocumento17 páginasCapsicum AnnumGloriana Cordero100% (1)
- Proyecto de Morfoanatomia Vegetal (La Hoja)Documento10 páginasProyecto de Morfoanatomia Vegetal (La Hoja)Jhonatan Lara LópezAún no hay calificaciones
- PRACTICA 6 y 7 de BotanicaDocumento8 páginasPRACTICA 6 y 7 de BotanicajamilAún no hay calificaciones
- Disertacion Hidrologia EvapotranspiracionDocumento2 páginasDisertacion Hidrologia EvapotranspiracionCesar Gutierrez NinahuamanAún no hay calificaciones
- Manual Cafetero Antiguo PDFDocumento401 páginasManual Cafetero Antiguo PDFcentro surcolombiano de investigación en café uscoAún no hay calificaciones
- Informe de Laboratorio Biologia.Documento23 páginasInforme de Laboratorio Biologia.sandra laverdeAún no hay calificaciones
- Clase2 Fisiologia Del ParasitismoDocumento33 páginasClase2 Fisiologia Del ParasitismoAnonymous 2HShWKOI100% (1)
- Aplicacion Foliar de Nutrientes - Retos y Limites en La Producción AgrícolaDocumento3 páginasAplicacion Foliar de Nutrientes - Retos y Limites en La Producción AgrícolaJessica Paola Tique ParraAún no hay calificaciones