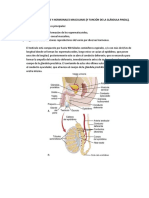
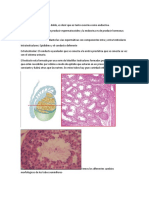
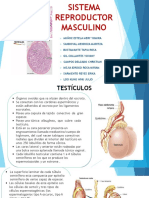
También podría gustarte
- Desarrollo HumanoembriologiaDocumento34 páginasDesarrollo HumanoembriologiaMaria Jose MolinaAún no hay calificaciones
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 páginasEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezAún no hay calificaciones
- GAMETOGENESISDocumento84 páginasGAMETOGENESISMery de Aguilar100% (1)
- OvogénesisDocumento3 páginasOvogénesisDenisse OvalleAún no hay calificaciones
- 1-RESUMEN GAMETOGENESISDocumento6 páginas1-RESUMEN GAMETOGENESISmateobeltri99Aún no hay calificaciones
- Gametogénesis: Formación de gametos masculinos y femeninosDocumento3 páginasGametogénesis: Formación de gametos masculinos y femeninosErik Lopez MamaniAún no hay calificaciones
- Ciclo Ovarico, Ovulación, Fecundación, Primera Semana, ImplantaciónDocumento10 páginasCiclo Ovarico, Ovulación, Fecundación, Primera Semana, ImplantaciónAstrid SofíaAún no hay calificaciones
- Gametogénesis y ciclo sexual.docxDocumento4 páginasGametogénesis y ciclo sexual.docxValentina BOLDRINIAún no hay calificaciones
- Resumen EmbriologíaDocumento4 páginasResumen Embriologíamonsesofia83Aún no hay calificaciones
- Resumen GametogenesisDocumento7 páginasResumen GametogenesisINSTITUTO MEDERIAún no hay calificaciones
- Espermatogénesis: Formación de EspermatozoidesDocumento9 páginasEspermatogénesis: Formación de EspermatozoidesJasminii Flor Llatas MontenegroAún no hay calificaciones
- Embriología GeneralDocumento23 páginasEmbriología Generalnicolas torresAún no hay calificaciones
- GAMETOGENESISDocumento6 páginasGAMETOGENESISCamilaAún no hay calificaciones
- Espermatogenesis - TrabajoDocumento12 páginasEspermatogenesis - TrabajoJanella Tineo MedinaAún no hay calificaciones
- Selem Channan Trabajo de FisiologíaDocumento7 páginasSelem Channan Trabajo de FisiologíaKenary GonzalezAún no hay calificaciones
- GametogenesisDocumento6 páginasGametogenesisRodriguez KarlaAún no hay calificaciones
- Fisiología de GametogénesisDocumento5 páginasFisiología de Gametogénesispier manuel cabezas armellonAún no hay calificaciones
- Game To GenesisDocumento36 páginasGame To Genesismaria ines rico zeferinoAún no hay calificaciones
- Gametognesis BiologiaDocumento15 páginasGametognesis BiologiaJosue RebolledoAún no hay calificaciones
- Gametogénesis, Ovulación e ImplantaciónDocumento31 páginasGametogénesis, Ovulación e ImplantaciónKeren Aquino BrunoAún no hay calificaciones
- Sistema ResumidoDocumento24 páginasSistema ResumidoJorge RamirezAún no hay calificaciones
- Gametogenesis M1.3Documento10 páginasGametogenesis M1.3Hector Efrain Garcia ChavezAún no hay calificaciones
- Embrio LOGIADocumento18 páginasEmbrio LOGIAGo Kids VpAún no hay calificaciones
- E SpermatogenesisDocumento6 páginasE SpermatogenesisThalía YanchapaxiAún no hay calificaciones
- En La Reproducción SexualDocumento2 páginasEn La Reproducción Sexualrosina paola siancas tineo100% (1)
- Proceso GametogénesisDocumento4 páginasProceso GametogénesisAlexander EztradaAún no hay calificaciones
- EMI MonografiaDocumento25 páginasEMI MonografiaAlejandro Mendoza SarmientoAún no hay calificaciones
- Funciones Reproductoras y Hormonales MasculinasDocumento10 páginasFunciones Reproductoras y Hormonales MasculinasMónica GonzálezAún no hay calificaciones
- Qué Es La GametogénesisDocumento3 páginasQué Es La GametogénesisRubenAún no hay calificaciones
- Biología Del Desarrollo IIDocumento67 páginasBiología Del Desarrollo IIMariaAún no hay calificaciones
- Anatomofisiologia ?Documento9 páginasAnatomofisiologia ?nirvanaojitoAún no hay calificaciones
- Resumen Embriologia LangmanDocumento13 páginasResumen Embriologia Langmanmanuel_aguirre_13Aún no hay calificaciones
- Clase 7 FecundaciónDocumento39 páginasClase 7 Fecundaciónjorge torricoAún no hay calificaciones
- Resumen Embirologia 2º SemestreDocumento9 páginasResumen Embirologia 2º SemestreDaniela RodríguezAún no hay calificaciones
- 2-Meiosis y GametogenesisDocumento48 páginas2-Meiosis y GametogenesisJesus Efren Villa GarciaAún no hay calificaciones
- Embriología: Desarrollo Ovulo Espermatozoide FecundaciónDocumento35 páginasEmbriología: Desarrollo Ovulo Espermatozoide FecundaciónLissetteAlexandraPerezAún no hay calificaciones
- Primera Semana de Desarrollo Moore PDFDocumento6 páginasPrimera Semana de Desarrollo Moore PDFAdrian FreudAún no hay calificaciones
- Resumen de Videos Parcial 1Documento65 páginasResumen de Videos Parcial 1Medicina BlogAún no hay calificaciones
- EMBRIOLOGIADocumento9 páginasEMBRIOLOGIAAlejandra MartinezAún no hay calificaciones
- GAMETOGENÉSISDocumento6 páginasGAMETOGENÉSISanon_757572911Aún no hay calificaciones
- Qué Son Los FosfolípidosDocumento33 páginasQué Son Los FosfolípidosNeftali Ascencio CastiblancoAún no hay calificaciones
- Division CelularDocumento51 páginasDivision CelularJosselin Jazmin Zanga QuispeAún no hay calificaciones
- Espermatogenesis y OvogenesisDocumento13 páginasEspermatogenesis y OvogenesisElvis Humantupa Estrada100% (3)
- La FecundaciónDocumento3 páginasLa FecundaciónjuberAún no hay calificaciones
- Espermatogenesis YfoliculogenesisDocumento9 páginasEspermatogenesis YfoliculogenesisJesus Gabriel Avila HernandezAún no hay calificaciones
- GametolgenesisDocumento18 páginasGametolgenesisFernando XochitecatlAún no hay calificaciones
- EmbriologíaDocumento24 páginasEmbriologíaJuan Andrés RestrepoAún no hay calificaciones
- Reproduccion Animal - 2Documento24 páginasReproduccion Animal - 2RUBEN MORALES BETANCURAún no hay calificaciones
- Sistema Reproductor Masculino (1) ORIGINALDocumento24 páginasSistema Reproductor Masculino (1) ORIGINALCristian CamposAún no hay calificaciones
- Cap 2 Embrio ResumenDocumento3 páginasCap 2 Embrio ResumenGABRIELA MONSERRAT MEJIA MONROYAún no hay calificaciones
- Gametogénesis y meiosis: La formación de gametosDocumento7 páginasGametogénesis y meiosis: La formación de gametosHeidy Juliana Ramírez GómezAún no hay calificaciones
- Marco Teórico: Generalidades Del Aparato Reproductor MasculinoDocumento18 páginasMarco Teórico: Generalidades Del Aparato Reproductor MasculinoAndrea IriasAún no hay calificaciones
- Primera Semana Del Desarrollo EmbrionarioDocumento33 páginasPrimera Semana Del Desarrollo EmbrionarioIsabella GomezAún no hay calificaciones
- Gametogenesis y FecundacionDocumento8 páginasGametogenesis y FecundacionCRISTIAMEAún no hay calificaciones
- Game To GenesisDocumento14 páginasGame To GenesisKaren ÜAún no hay calificaciones
- FECUNDACIONDocumento31 páginasFECUNDACIONTonyRequejoNúñezAún no hay calificaciones
- Biologia de La Concepcion Aparato Reproductor (Autoguardado)Documento16 páginasBiologia de La Concepcion Aparato Reproductor (Autoguardado)Eliana Velasco100% (1)
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Analizando las características del Despotismo IlustradoDocumento3 páginasAnalizando las características del Despotismo Ilustradosfs100% (4)
- El arte como construcción humana a través del tiempoDocumento14 páginasEl arte como construcción humana a través del tiempoFeer CabelloAún no hay calificaciones
- Comunicación I ResumenDocumento26 páginasComunicación I ResumenBrunoAún no hay calificaciones
- Jubert, Roxane. Continuación y ReconstrucciónDocumento7 páginasJubert, Roxane. Continuación y ReconstrucciónBrunoAún no hay calificaciones
- Resumen Final Comunicación Arfuch (Ahora Devalle) (Cursada 2015)Documento24 páginasResumen Final Comunicación Arfuch (Ahora Devalle) (Cursada 2015)Meme Ceresole67% (3)
- CRONOGRAMA COM1 1er CUATRIMESTRE 2022Documento3 páginasCRONOGRAMA COM1 1er CUATRIMESTRE 2022BrunoAún no hay calificaciones
- Comunicación I ResumenDocumento2 páginasComunicación I ResumenBrunoAún no hay calificaciones
- Formas Obtención 2020 ColadaDocumento21 páginasFormas Obtención 2020 ColadaBrunoAún no hay calificaciones
- Eco, UDocumento3 páginasEco, UBrunoAún no hay calificaciones
- Comunicación I ResumenDocumento3 páginasComunicación I ResumenBrunoAún no hay calificaciones
- Lazo, Alfonso. Revolución RusaDocumento9 páginasLazo, Alfonso. Revolución RusaBrunoAún no hay calificaciones
- Instrumentación Mecanizada IDocumento7 páginasInstrumentación Mecanizada IBrunoAún no hay calificaciones
- Gripe vs ResfriadoDocumento6 páginasGripe vs ResfriadoBrunoAún no hay calificaciones
- Chernevich, Elena. Constructor-Publicitario. Maiakovsky-RodchenkoDocumento6 páginasChernevich, Elena. Constructor-Publicitario. Maiakovsky-RodchenkoBrunoAún no hay calificaciones
- Maq CorazDocumento1 páginaMaq CorazDaigoro LoveraAún no hay calificaciones
- Cuentos Con Jesus 5 PDFDocumento42 páginasCuentos Con Jesus 5 PDFLina Fernanda Pino50% (6)
- Resumen 2do Parcial HistoriaDocumento6 páginasResumen 2do Parcial HistoriaBrunoAún no hay calificaciones
- Dios Nos Creo y Nos CuidaDocumento29 páginasDios Nos Creo y Nos CuidaBrunoAún no hay calificaciones
- Clase 2 Impresiones y ModelosDocumento4 páginasClase 2 Impresiones y ModelosBrunoAún no hay calificaciones
- Clase 3. Articuladores, Inducción y MontajeDocumento6 páginasClase 3. Articuladores, Inducción y MontajeBrunoAún no hay calificaciones
- Cuadro ComparativoDocumento1 páginaCuadro ComparativoBrunoAún no hay calificaciones
- Clase 2 Impresiones y ModelosDocumento4 páginasClase 2 Impresiones y ModelosBrunoAún no hay calificaciones
- Untitled DocumentDocumento1 páginaUntitled DocumentBrunoAún no hay calificaciones
- CLASE 2 Corrección de ModelosDocumento2 páginasCLASE 2 Corrección de ModelosBruno Aurelio Huallan PadillaAún no hay calificaciones
- GENERALIDADESDocumento19 páginasGENERALIDADESBrunoAún no hay calificaciones
- Espacio y Representación - Grupo 2Documento5 páginasEspacio y Representación - Grupo 2BrunoAún no hay calificaciones
- Tipografía y expresión ciclo 2021Documento1 páginaTipografía y expresión ciclo 2021BrunoAún no hay calificaciones
- Pautas Neurofisiología 2020Documento1 páginaPautas Neurofisiología 2020BrunoAún no hay calificaciones
- PinterestDocumento8 páginasPinterestBrunoAún no hay calificaciones
- ConceptoDocumento1 páginaConceptoBrunoAún no hay calificaciones
- Enfermedades clostridiales bovinas: causas, síntomas y prevenciónDocumento95 páginasEnfermedades clostridiales bovinas: causas, síntomas y prevenciónDilmer GuzmanAún no hay calificaciones
- Cereales Andinos (Monografia)Documento20 páginasCereales Andinos (Monografia)elizitasol100% (7)
- M203 AnatomíaDocumento34 páginasM203 AnatomíaSebas VidalAún no hay calificaciones
- 01 BoliviaDocumento84 páginas01 BoliviaPaolita Pinzón ArangoAún no hay calificaciones
- Informe 8Documento5 páginasInforme 8Carlos Andres GAún no hay calificaciones
- 401 Alteraciones EquilibrioDocumento7 páginas401 Alteraciones EquilibrioLisset AdasmeAún no hay calificaciones
- Concepto y Ambito de La PersonalidadDocumento41 páginasConcepto y Ambito de La PersonalidadMoises Nomeli Molina LopezAún no hay calificaciones
- Ud 4 Técnicas SiembraDocumento5 páginasUd 4 Técnicas SiembraLiliana LinoAún no hay calificaciones
- Actividad 4 - Cuadro ComparativoDocumento5 páginasActividad 4 - Cuadro ComparativoLina FernandezAún no hay calificaciones
- Hipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasDocumento10 páginasHipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasFelipe Javier Melis ArcosAún no hay calificaciones
- 36 Puntos Bubishi OkinawaDocumento40 páginas36 Puntos Bubishi OkinawaGuillermo Daniel Durán100% (1)
- Herencias genéticas y árboles genealógicosDocumento8 páginasHerencias genéticas y árboles genealógicosHyllary BorjaAún no hay calificaciones
- Fase 2-Locomoción Humana - Greys HernandezDocumento21 páginasFase 2-Locomoción Humana - Greys HernandezGrace Hernandez Cervantes100% (1)
- Influencias en El DesarrolloDocumento37 páginasInfluencias en El DesarrolloHugo Carmona JasmeAún no hay calificaciones
- Distribución y características de tepezcuintles y seretes en MéxicoDocumento3 páginasDistribución y características de tepezcuintles y seretes en MéxicoNathan Ulises Reyes JimenezAún no hay calificaciones
- Aap C05 U1 EvPro 1a5Documento2 páginasAap C05 U1 EvPro 1a5Lizeth CubillosAún no hay calificaciones
- Estudio de Cinco Cepas Fungosas en GranadillaDocumento29 páginasEstudio de Cinco Cepas Fungosas en Granadillamanuelborgefenix1362Aún no hay calificaciones
- Masaje DescontracturanteDocumento2 páginasMasaje DescontracturanteFamilia cristiana unidaAún no hay calificaciones
- Trabajo BiofisicaDocumento8 páginasTrabajo Biofisicazuluagaja0075918Aún no hay calificaciones
- Examlab 112690175 201220221835Documento4 páginasExamlab 112690175 201220221835Marcela UmañaAún no hay calificaciones
- Guia de Estudio para Biologia PDFDocumento37 páginasGuia de Estudio para Biologia PDFMari Ad LaeAún no hay calificaciones
- Ecologia de Las PoblacionesDocumento4 páginasEcologia de Las PoblacionesS3MAXAún no hay calificaciones
- MyE. Tema 2 - La Perspectiva Motivacional, ¿Por Qué Hacemos Lo Que HacemosDocumento8 páginasMyE. Tema 2 - La Perspectiva Motivacional, ¿Por Qué Hacemos Lo Que HacemosCARLOTA CALLES PÉREZAún no hay calificaciones
- CanidaeDocumento6 páginasCanidaeRuben Villamizar FlorezAún no hay calificaciones
- Procesamiento visual y proyecciones subcorticalesDocumento22 páginasProcesamiento visual y proyecciones subcorticalesYuliana FrancoAún no hay calificaciones
- CAPÍTULO 2 LavisiónDocumento27 páginasCAPÍTULO 2 LavisiónAgueda AnabelAún no hay calificaciones
- PÁNCREASDocumento10 páginasPÁNCREASstefany ortegaAún no hay calificaciones
- Tejido Epitelial ProyectoDocumento3 páginasTejido Epitelial ProyectoArturo EspinozaAún no hay calificaciones
- Prevención MARSI y LESCAH: Enfoque inicial para lesiones cutáneasDocumento51 páginasPrevención MARSI y LESCAH: Enfoque inicial para lesiones cutáneasSantiago Estrada100% (1)
- Sonda Nasogástrica EquinosDocumento14 páginasSonda Nasogástrica EquinosJohacvAún no hay calificaciones