También podría gustarte
- Infante Con Impedimentos Cognitivos y MotoresDocumento24 páginasInfante Con Impedimentos Cognitivos y MotoresDaniel Lopez100% (4)
- Procesamiento de La Actividad MotoraDocumento38 páginasProcesamiento de La Actividad MotoraGian Marco villena67% (3)
- VÍAS MOTORAS y SENSITIVASDocumento5 páginasVÍAS MOTORAS y SENSITIVASSabrina Rivero100% (5)
- Actividad ReflejaDocumento71 páginasActividad ReflejaTito Valdés Córdova67% (12)
- Facilitación Neuromuscular PropioceptivaDocumento58 páginasFacilitación Neuromuscular Propioceptivarpalaciosf100% (3)
- Lesion Medular TripticoDocumento1 páginaLesion Medular TripticoRonald Alfredo Llallico Echevarria100% (1)
- Corteza motora: estructura y funciones principalesDocumento17 páginasCorteza motora: estructura y funciones principalesBryan JuarezAún no hay calificaciones
- Rehabilitación en Alteraciones de La SensibilidadDocumento23 páginasRehabilitación en Alteraciones de La SensibilidadYully Chaparro Jimenez100% (1)
- Ficha de Evaluación Fisioterapéutica de Parálisis Cerebral InfantilDocumento3 páginasFicha de Evaluación Fisioterapéutica de Parálisis Cerebral InfantilEvelyn Rojas80% (5)
- Evaluacion KinesicaDocumento2 páginasEvaluacion KinesicasebaAún no hay calificaciones
- SCIM Inareps Lesion Medular Anexo 3Documento4 páginasSCIM Inareps Lesion Medular Anexo 3Verónica SenatoreAún no hay calificaciones
- Interacciones de Las Células Con Los Materiales ExtracelularesDocumento7 páginasInteracciones de Las Células Con Los Materiales ExtracelularesFlor CarreraAún no hay calificaciones
- DESARROLLO Motor Atipico TipicoDocumento29 páginasDESARROLLO Motor Atipico TipicoGonzalo Aguayo Alcayaga50% (2)
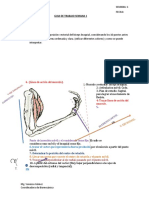
- Guia 2 AcabadaDocumento5 páginasGuia 2 AcabadaJotaro JoestarAún no hay calificaciones
- Jebsen Taylor FichaDocumento1 páginaJebsen Taylor FichaGerman BuyattiAún no hay calificaciones
- Curso Estimulación TempranaDocumento99 páginasCurso Estimulación TempranaValeria Gallardo Vargas100% (3)
- SinapsisDocumento43 páginasSinapsisValeria EnrriquezAún no hay calificaciones
- Contracciones muscularesDocumento16 páginasContracciones muscularesRoggert CrodovaAún no hay calificaciones
- Biomecanica de La ColumnaDocumento35 páginasBiomecanica de La ColumnaJeff VillaplanaAún no hay calificaciones
- Control Postural AnormalDocumento28 páginasControl Postural AnormalAndrés MardonesAún no hay calificaciones
- Sistema PropioceptivoDocumento2 páginasSistema PropioceptivoGuille HuertaAún no hay calificaciones
- Áreas de Brodmann y sus funcionesDocumento9 páginasÁreas de Brodmann y sus funcionesGénesis SánchezAún no hay calificaciones
- Temas de Estatica y Dinamica PDFDocumento48 páginasTemas de Estatica y Dinamica PDFEmilio Diaz RodriguezAún no hay calificaciones
- Mapa Conseptual de Memoria y OlvidoDocumento1 páginaMapa Conseptual de Memoria y OlvidoNeider LopezAún no hay calificaciones
- Método Rood PDFDocumento2 páginasMétodo Rood PDFLeo_Ram98100% (1)
- Modelo Cinesiologico, Patocinesiologico, CinesipatologicoDocumento4 páginasModelo Cinesiologico, Patocinesiologico, CinesipatologicoANGIE PESEROSAún no hay calificaciones
- Triada Infeliz de RodillaDocumento9 páginasTriada Infeliz de RodillaCarIto Vergara0% (1)
- Evaluación Básica Del Paciente Neurológico ListoDocumento13 páginasEvaluación Básica Del Paciente Neurológico ListoFrancisco Javier Luza Ramos100% (1)
- Enfermedad de HuntingtonDocumento16 páginasEnfermedad de HuntingtonMelissa SilvaAún no hay calificaciones
- Principios Básicos de ElectroterapiaDocumento44 páginasPrincipios Básicos de ElectroterapiaAlejandro Sánchez PriegoAún no hay calificaciones
- Proceso de Mielinización o MielyogenesisDocumento5 páginasProceso de Mielinización o MielyogenesisEugenia Carabajal0% (1)
- Escala de BarthelDocumento7 páginasEscala de BarthelJames Castillo GasteloAún no hay calificaciones
- Vias AscendentesDocumento24 páginasVias AscendentesAnderson GilAún no hay calificaciones
- Método PhelpsDocumento11 páginasMétodo PhelpsJuanita MonterrosoAún no hay calificaciones
- Modelos Teóricos y Prácticos de RehabilitaciónDocumento6 páginasModelos Teóricos y Prácticos de RehabilitaciónAdriana Saenz AlvaresAún no hay calificaciones
- Prueba de AdamsDocumento12 páginasPrueba de Adamstonni marchena100% (1)
- Control PosturalDocumento72 páginasControl PosturalZandraTC100% (1)
- SinergiasDocumento3 páginasSinergiasCarlos Alvarado0% (1)
- Presentacion Power Point de Control Motor Martes 140818Documento14 páginasPresentacion Power Point de Control Motor Martes 140818Adriana Lucia Martinez CastillaAún no hay calificaciones
- Neuro 2 PDFDocumento4 páginasNeuro 2 PDFRamon Vicente Linares PerezAún no hay calificaciones
- Modelo de Sharman para el diagnóstico y tratamiento de alteraciones del movimientoDocumento1 páginaModelo de Sharman para el diagnóstico y tratamiento de alteraciones del movimientoSheyla EscuderoAún no hay calificaciones
- Reacciones Neuromotrices.Documento9 páginasReacciones Neuromotrices.Paula Andrea GómezAún no hay calificaciones
- Neuroplasticidad Expo NataliaDocumento32 páginasNeuroplasticidad Expo NataliaNATALIA BRONSTEINAún no hay calificaciones
- Teoria de La Seleccion de Grupos NeuronalesDocumento4 páginasTeoria de La Seleccion de Grupos NeuronalesDorian Wilfredo Juli AnchapuriAún no hay calificaciones
- Casos ClínicosDocumento17 páginasCasos ClínicosmarcelaAún no hay calificaciones
- Biomecánica Del HuesoDocumento22 páginasBiomecánica Del HuesoJos� Leonardo M�ndez PossoAún no hay calificaciones
- Caso Clínico - HombroDocumento2 páginasCaso Clínico - HombroKaren Benita Mena PastranaAún no hay calificaciones
- Resultado de aprendizaje en neurodesarrolloDocumento55 páginasResultado de aprendizaje en neurodesarrolloAdolfo CarrascoAún no hay calificaciones
- Tecnicas de Facilitacion e InhibicionDocumento28 páginasTecnicas de Facilitacion e InhibicionSimon Nuñez Urquieta100% (3)
- Tratamiento fisioterapia cuadriparesiaDocumento4 páginasTratamiento fisioterapia cuadriparesianegrita84100% (4)
- Cuadro de ReflejosDocumento5 páginasCuadro de ReflejosMedicina Fisica Rehabilitacion Integral33% (3)
- Método de Phelps para PCDocumento31 páginasMétodo de Phelps para PCJade Irem Velazquez100% (1)
- BOBATH de Sentado A Parado - En.esDocumento31 páginasBOBATH de Sentado A Parado - En.esLizeth ArceAún no hay calificaciones
- Evaluación cognitiva MMSEDocumento5 páginasEvaluación cognitiva MMSEalaneduardo14100% (1)
- Aprendizaje motor y recuperación funcionalDocumento17 páginasAprendizaje motor y recuperación funcionalLaura Fleitas Bóveda67% (3)
- Motoneurona Superior e InferiorDocumento35 páginasMotoneurona Superior e InferiorJohanna Galvis100% (2)
- Ferreres Modulo II BDocumento56 páginasFerreres Modulo II BDiego BilbaoAún no hay calificaciones
- Apunte Sistema MotorDocumento6 páginasApunte Sistema MotorPABLO EMILIO PINO RIVASAún no hay calificaciones
- Fundamentos de Fisiologia Del EjercicioDocumento31 páginasFundamentos de Fisiologia Del EjercicioAlvaro Monsalve100% (1)
- wuolah-free-TEMA 3. SISTEMA NERVIOSO. SISTEMA MOTOR Y SU CONTROLDocumento9 páginaswuolah-free-TEMA 3. SISTEMA NERVIOSO. SISTEMA MOTOR Y SU CONTROLelcr4ck21.1Aún no hay calificaciones
- Unidad motora y reflejos espinalesDocumento12 páginasUnidad motora y reflejos espinalesJessi MenezesAún no hay calificaciones
- (7 Fisiología Muscular) PDFDocumento12 páginas(7 Fisiología Muscular) PDFOrlando J SierraAún no hay calificaciones
- (Capítulo F3. Control Suprasegmentario Integración Medular)Documento14 páginas(Capítulo F3. Control Suprasegmentario Integración Medular)Orlando J SierraAún no hay calificaciones
- (Capítulo F2. Reflejos Medulares II)Documento16 páginas(Capítulo F2. Reflejos Medulares II)Orlando J SierraAún no hay calificaciones
- (Capítulo F1. El Sistema Nervioso. Reflejos Medulares I)Documento16 páginas(Capítulo F1. El Sistema Nervioso. Reflejos Medulares I)Orlando J SierraAún no hay calificaciones
- 9control Neural Del Movimiento - Luis Aguilar PDFDocumento30 páginas9control Neural Del Movimiento - Luis Aguilar PDFMaria Cristina Nates HuertasAún no hay calificaciones
- Anatomía SNDocumento148 páginasAnatomía SNJesica LegazpiaAún no hay calificaciones
- El Sistema Nervioso HumanoDocumento4 páginasEl Sistema Nervioso HumanoLorenBarckleyAún no hay calificaciones
- Unidad 5 Inteligencia ArtificialDocumento21 páginasUnidad 5 Inteligencia ArtificialPedro ChavarriaAún no hay calificaciones
- Compendio Lecturas 2021-0 - Pages - DeletedDocumento10 páginasCompendio Lecturas 2021-0 - Pages - DeletedAlessandraAún no hay calificaciones
- Temario de GeriatríaDocumento13 páginasTemario de GeriatríaUNIAún no hay calificaciones
- Apunte C - El Aula Curiosa y El AprendizajeDocumento7 páginasApunte C - El Aula Curiosa y El AprendizajeMariela LedesmaAún no hay calificaciones
- REVISTA DE INTELIGENCIA - Desarollo de La Inteligencia PDFDocumento5 páginasREVISTA DE INTELIGENCIA - Desarollo de La Inteligencia PDFMariana SánchezAún no hay calificaciones
- Marco TeoricoDocumento2 páginasMarco TeoricoGloria Liliana Cañón Arango100% (1)
- La Coordinacion Motriz A Distancia (Argemi Barrios)Documento4 páginasLa Coordinacion Motriz A Distancia (Argemi Barrios)Juan Sebastian Ortiz AyalaAún no hay calificaciones
- Oral Health and Healthy Chewing For Healthy Cognitive Ageing - A Comprehensive Narrative ReviewDocumento10 páginasOral Health and Healthy Chewing For Healthy Cognitive Ageing - A Comprehensive Narrative ReviewAMELIA PAZ LETELIER AGUIRREAún no hay calificaciones
- Es El Estómago El Segundo CerebroDocumento9 páginasEs El Estómago El Segundo CerebroLuciano AlcaláAún no hay calificaciones
- Parcial - Escenario 4 - Comportamiento Del Consumidor - 202060-b2 - b03Documento10 páginasParcial - Escenario 4 - Comportamiento Del Consumidor - 202060-b2 - b03Laidy MorenoAún no hay calificaciones
- Aparatos y sistemas del cuerpo humanoDocumento23 páginasAparatos y sistemas del cuerpo humanoRocio Galindo GarciaAún no hay calificaciones
- Los Sistemas EfectoresDocumento13 páginasLos Sistemas EfectoresCarola AilínAún no hay calificaciones
- La plasticidad cerebral, neuronas espejo y poda neuronal en relación con el aprendizajeDocumento3 páginasLa plasticidad cerebral, neuronas espejo y poda neuronal en relación con el aprendizajeJoan Alexander Loli JaénAún no hay calificaciones
- TELOS 81 - Cuadernos de Comunicacion e InnovacionDocumento191 páginasTELOS 81 - Cuadernos de Comunicacion e InnovacionPilu LlambiasAún no hay calificaciones
- Neuronas y sus estructurasDocumento6 páginasNeuronas y sus estructurasfelipe jaraAún no hay calificaciones
- Resumen Capitulo 55Documento8 páginasResumen Capitulo 55Rashel MoscosoAún no hay calificaciones
- Historia de Las Neurociencias de La ConductaDocumento4 páginasHistoria de Las Neurociencias de La ConductaLaura SilvaAún no hay calificaciones
- Neurociencia y ConductaDocumento17 páginasNeurociencia y ConductaShalini Quiroz BecerraAún no hay calificaciones
- Anna Lucia Campos, PY 1, Neurociencias Aprendizaje y NeuroeducaciónDocumento35 páginasAnna Lucia Campos, PY 1, Neurociencias Aprendizaje y NeuroeducaciónserpaolaAún no hay calificaciones
- Tálamo ÓpticoDocumento4 páginasTálamo ÓpticoMatias CostanzoAún no hay calificaciones
- AA1 Neuronas y Potenciales de MembranaDocumento4 páginasAA1 Neuronas y Potenciales de MembranaFernando CadenaAún no hay calificaciones
- Anatomía y Fisiología Del Sistema Nervioso TAREA 3Documento9 páginasAnatomía y Fisiología Del Sistema Nervioso TAREA 3bohane santos0% (1)
- Portafolio 1 de Histologia y AnatomiaDocumento8 páginasPortafolio 1 de Histologia y AnatomiaItzel LeyvaAún no hay calificaciones
- FisiologadelanarizDocumento6 páginasFisiologadelanarizroAún no hay calificaciones
- Nocicepcion y DolorDocumento13 páginasNocicepcion y DolorLucia SanchezAún no hay calificaciones
- Tinciones para Tejido Nervioso (Eq6)Documento7 páginasTinciones para Tejido Nervioso (Eq6)fer.est.lop10Aún no hay calificaciones
- PsicofarmacoDocumento531 páginasPsicofarmacoTHAIS GONZALES INDACOCHEAAún no hay calificaciones