También podría gustarte
- Resultados Compensar PDFDocumento2 páginasResultados Compensar PDFSonia Henao Valencia100% (1)
- Ciclo de KrebsDocumento12 páginasCiclo de KrebsGallardo RichardAún no hay calificaciones
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsMiguel Angel Rodas Herrera100% (2)
- Curva SDocumento5 páginasCurva Sadonis100% (1)
- Manual Tecnico Descriptivo Protocolo de Comunicacion RIGAZSADocumento91 páginasManual Tecnico Descriptivo Protocolo de Comunicacion RIGAZSAsammary00Aún no hay calificaciones
- Metabolismo CatabolismoDocumento33 páginasMetabolismo CatabolismoFrancis A. Batarse0% (1)
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsLN Brenda Janeth Beltrán DíazAún no hay calificaciones
- El Catabolismo y Anabolismo 2aDocumento17 páginasEl Catabolismo y Anabolismo 2aRuthm GhAún no hay calificaciones
- Resumen de Rutas Metabólicas BCMDocumento52 páginasResumen de Rutas Metabólicas BCMLuana HuertasAún no hay calificaciones
- CATABOLISMODocumento8 páginasCATABOLISMOMerry Peralta VillanuevaAún no hay calificaciones
- Ciclo de KrebsDocumento19 páginasCiclo de KrebsWikii Wikoso0% (1)
- Camara de ContactoDocumento6 páginasCamara de Contactoadonis100% (3)
- Respiracion Celular IIDocumento31 páginasRespiracion Celular IIDelvis SánchezAún no hay calificaciones
- Planeación de Requerimiento de Los Materiales. ADMINISTRACIÓN DE OPERACIONES IIDocumento16 páginasPlaneación de Requerimiento de Los Materiales. ADMINISTRACIÓN DE OPERACIONES IIZULEM GUTIERREZAún no hay calificaciones
- CatabolismoDocumento7 páginasCatabolismocarolAún no hay calificaciones
- CatabolismoDocumento8 páginasCatabolismoEiner SanjuaneloAún no hay calificaciones
- Convergencia y Diversidad en El CatabolismoDocumento5 páginasConvergencia y Diversidad en El CatabolismojanansayaAún no hay calificaciones
- El CatabolismoDocumento3 páginasEl CatabolismoLeonardo FernandezAún no hay calificaciones
- Relación Aminoácidos Con La Glucolisis y El Ciclo de KrebsDocumento5 páginasRelación Aminoácidos Con La Glucolisis y El Ciclo de KrebsMaria Fernanda Meza TapiaAún no hay calificaciones
- Ciclo KrebsDocumento13 páginasCiclo KrebsAlex Jimeno0% (1)
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsTania Lisset Oblea MejiaAún no hay calificaciones
- CUESTIONARIODocumento8 páginasCUESTIONARIOJuan Jose Baros AyalaAún no hay calificaciones
- Bioquimica MonografiaDocumento15 páginasBioquimica MonografiaCarolay ParedesAún no hay calificaciones
- Unidad 3Documento9 páginasUnidad 3daniel J Alejos CAún no hay calificaciones
- Equipo 10, Tema de Exposición.Documento24 páginasEquipo 10, Tema de Exposición.Fernanda IdkAún no hay calificaciones
- LipogénesisDocumento16 páginasLipogénesisgabycita910% (1)
- Introducción: Ciclo de KrebsDocumento5 páginasIntroducción: Ciclo de KrebsJuan David Losada RodriguezAún no hay calificaciones
- Colegio Adventista PimentelDocumento26 páginasColegio Adventista PimentelCamila RubioAún no hay calificaciones
- Tema 14 Ruta Ciclo de Krebs. Autor Alexander Sanchez Lamas.Documento19 páginasTema 14 Ruta Ciclo de Krebs. Autor Alexander Sanchez Lamas.Alexander SanchezAún no hay calificaciones
- Catabolismo PDFDocumento2 páginasCatabolismo PDFedgarAún no hay calificaciones
- Metabolismo Bioquimica PDFDocumento8 páginasMetabolismo Bioquimica PDFcarlos prudAún no hay calificaciones
- Ciclo de KrebsDocumento6 páginasCiclo de KrebsJose Luis Arroyo Hernandez0% (1)
- DunyDocumento9 páginasDunyMarisolAún no hay calificaciones
- Bioquimica - Diferencias de Celula Animal y VegetalDocumento5 páginasBioquimica - Diferencias de Celula Animal y VegetalRanthal Rivadeneyra CastroAún no hay calificaciones
- Quimica Tema 3Documento4 páginasQuimica Tema 3NandyAún no hay calificaciones
- CATABOLISMO (Aerobio y Anaerobio)Documento66 páginasCATABOLISMO (Aerobio y Anaerobio)Elizabetн Essel Cordilia Middleford50% (2)
- Respiración CelularDocumento3 páginasRespiración CelularAlexia NavarroAún no hay calificaciones
- Ciclo de Krebs, Cadena Respiratoria, Balance y Via de Las Pentosas. 2023Documento30 páginasCiclo de Krebs, Cadena Respiratoria, Balance y Via de Las Pentosas. 2023Lara VilteAún no hay calificaciones
- Modulo Metabolismo-TallerDocumento14 páginasModulo Metabolismo-TallerMafe BenitezAún no hay calificaciones
- Ciclo de Krebs Seminario 2dauDocumento8 páginasCiclo de Krebs Seminario 2dauDanixaKristelBarriosUcañayAún no hay calificaciones
- Ciclo de Krebs - Exposición de F. de Micro y BioDocumento29 páginasCiclo de Krebs - Exposición de F. de Micro y BioMaria Jose QuirozAún no hay calificaciones
- Ciclo de Krebs BasicoDocumento9 páginasCiclo de Krebs BasicoJavier AranaAún no hay calificaciones
- Objetivo 2 Ciclo de KrebsDocumento4 páginasObjetivo 2 Ciclo de KrebsKalii ZamoraAún no hay calificaciones
- Ciclo de KrebsDocumento10 páginasCiclo de KrebsEliannys Arregoces BritoAún no hay calificaciones
- El Ciclo de KrebsDocumento8 páginasEl Ciclo de KrebsTlacaélel Rafael Cáceres Santa CruzAún no hay calificaciones
- Metabolismo Gluco Kbres MedicinaDocumento40 páginasMetabolismo Gluco Kbres MedicinaLuis eduardo La torre AdrianzénAún no hay calificaciones
- Resumen 12 Ciclo de KrebsDocumento12 páginasResumen 12 Ciclo de KrebsMariam Cabellos SAún no hay calificaciones
- Tema 4Documento6 páginasTema 4Maria GarciaAún no hay calificaciones
- Metabolismo Catabolismo y AnabolismoDocumento25 páginasMetabolismo Catabolismo y AnabolismoIngrid MathieuAún no hay calificaciones
- Piruvato DeshidrogenasaDocumento7 páginasPiruvato DeshidrogenasaRoberth Daniel Torres GutierrezAún no hay calificaciones
- Catabolismo de Lipidos en VegetalesDocumento32 páginasCatabolismo de Lipidos en VegetalesgabyvargasAún no hay calificaciones
- Respiración Celular 2021Documento29 páginasRespiración Celular 202110HorasAún no hay calificaciones
- Metabolismo y Respiracion CelularDocumento25 páginasMetabolismo y Respiracion CelularValeria GuerraAún no hay calificaciones
- Ciclo de Krebs. Grupo 4. Bioquímica II.Documento11 páginasCiclo de Krebs. Grupo 4. Bioquímica II.Zuly AGomezAún no hay calificaciones
- Catabolismo Y Anabolismo: García Hernández Ruth María. Lic. Cultura Física y DeportesDocumento16 páginasCatabolismo Y Anabolismo: García Hernández Ruth María. Lic. Cultura Física y Deporteslina garciaAún no hay calificaciones
- El Ciclo de KrebsDocumento3 páginasEl Ciclo de KrebsɆsƬeŁa尺Aún no hay calificaciones
- Ciclo de KrebsDocumento7 páginasCiclo de KrebsRenzo GonzalezAún no hay calificaciones
- Acetil-CoA, Ciclo de KrebsDocumento12 páginasAcetil-CoA, Ciclo de KrebsClauMcClauAún no hay calificaciones
- Ciclo de Krebs y Ciclo de Urea ExpoDocumento14 páginasCiclo de Krebs y Ciclo de Urea Expomay_street_von100% (1)
- Metabolitos IntermediariosDocumento3 páginasMetabolitos IntermediariosFelipe ChimborazoAún no hay calificaciones
- Taller de Bioquimica - MetabolismoDocumento14 páginasTaller de Bioquimica - MetabolismoValeria Andrea Carrillo Contreras100% (1)
- Laura BioquimicaDocumento19 páginasLaura BioquimicaLauren AmayaAún no hay calificaciones
- Ciclo de KlebsDocumento9 páginasCiclo de KlebsIram Fernando Oleta CruzAún no hay calificaciones
- TEMA 6 2º Bach METABOLISMODocumento21 páginasTEMA 6 2º Bach METABOLISMOMarina Ramos GarciaAún no hay calificaciones
- Informe 394-2022-DGPIGA-DGEIADocumento9 páginasInforme 394-2022-DGPIGA-DGEIAadonisAún no hay calificaciones
- Un Muro de ContenciónDocumento2 páginasUn Muro de ContenciónadonisAún no hay calificaciones
- Vibrador de ConcretoDocumento6 páginasVibrador de ConcretoadonisAún no hay calificaciones
- Agua BlandaDocumento7 páginasAgua BlandaadonisAún no hay calificaciones
- Instalacionese 110214141210 Phpapp02Documento84 páginasInstalacionese 110214141210 Phpapp02Eduardo FonsecaAún no hay calificaciones
- DesagüeDocumento5 páginasDesagüeadonisAún no hay calificaciones
- Tractor Sobre OrugasDocumento7 páginasTractor Sobre OrugasadonisAún no hay calificaciones
- Los GeosinteticosDocumento15 páginasLos GeosinteticosadonisAún no hay calificaciones
- Diseño de DesarenadorDocumento9 páginasDiseño de DesarenadoradonisAún no hay calificaciones
- ProyectosDocumento3 páginasProyectosadonisAún no hay calificaciones
- Planta de Tratamiento de Agua PotableDocumento6 páginasPlanta de Tratamiento de Agua PotableadonisAún no hay calificaciones
- Músculos de La Cara y Su FunciónDocumento6 páginasMúsculos de La Cara y Su Funciónramiro mercadoAún no hay calificaciones
- CanticoDocumento162 páginasCanticololoAún no hay calificaciones
- CheetttosDocumento1 páginaCheetttosdukegarrikAún no hay calificaciones
- Mec Flu IIM350 Lista de Ejercicios II Semestre I 2020Documento8 páginasMec Flu IIM350 Lista de Ejercicios II Semestre I 2020pedroAún no hay calificaciones
- Tarea 1 Etica y LegislaciónDocumento2 páginasTarea 1 Etica y LegislaciónLordmef Antonio LoretoAún no hay calificaciones
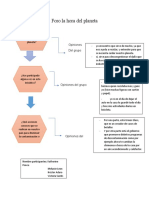
- Foro Del Planeta EsquemaDocumento1 páginaForo Del Planeta Esquemakatherine happyAún no hay calificaciones
- Acondicionamiento en VeranoDocumento25 páginasAcondicionamiento en VeranoLuisReyesMolinaAún no hay calificaciones
- Trabajo Final Implementacion Factura Electronica S.VDocumento23 páginasTrabajo Final Implementacion Factura Electronica S.VIsabel Cristina Huertas GarciaAún no hay calificaciones
- Propuesta de Juegos y Deportes Con Implementos Como Contenido Alternativo en El Área de Educación FísicaDocumento5 páginasPropuesta de Juegos y Deportes Con Implementos Como Contenido Alternativo en El Área de Educación FísicaluisAún no hay calificaciones
- Consagracion Sagrat Cor Torres I Bages CASCAT A5 PDFDocumento4 páginasConsagracion Sagrat Cor Torres I Bages CASCAT A5 PDFjoseluisvivesAún no hay calificaciones
- Necocli TrabajoDocumento8 páginasNecocli TrabajoAndrea MartinezAún no hay calificaciones
- Alteraciones Morfológicas en La Lesión CelularDocumento11 páginasAlteraciones Morfológicas en La Lesión CelularFlor FLorAún no hay calificaciones
- Guia de Tercero Diferenciado Metodos de Formacion de SalesDocumento5 páginasGuia de Tercero Diferenciado Metodos de Formacion de SalesHarold BustamanteAún no hay calificaciones
- 4° Año-Teoría 20Documento2 páginas4° Año-Teoría 20Jackson López LaviAún no hay calificaciones
- Cuencas SedimentariasDocumento32 páginasCuencas SedimentariasRonaldJoséCuencaRoa100% (1)
- Semana 5 NeuropsicologiaDocumento9 páginasSemana 5 NeuropsicologiaVictor Santos SAún no hay calificaciones
- Py Css PosterDocumento2 páginasPy Css PosterzeppvargasAún no hay calificaciones
- Practica Gestion Abastecimiento1ER PARCIALDocumento3 páginasPractica Gestion Abastecimiento1ER PARCIALFabiola PesantesAún no hay calificaciones
- Indroduccion A Proyecto de Licor de MielDocumento24 páginasIndroduccion A Proyecto de Licor de MielErick castellonAún no hay calificaciones
- Resumen - Mecanismos de EndurecimientoDocumento2 páginasResumen - Mecanismos de EndurecimientoJuan Wilfredo Hernandez DiazAún no hay calificaciones
- Plan de Clase Etica 11° Agosto 31Documento3 páginasPlan de Clase Etica 11° Agosto 31Pedro Pablo Polo FlorezAún no hay calificaciones
- Informe Expocitorio: Los Materiales Sólidos y Líquidos, Siendo La Proliferación de Basuras Uno deDocumento4 páginasInforme Expocitorio: Los Materiales Sólidos y Líquidos, Siendo La Proliferación de Basuras Uno deJuan Luis OrsanAún no hay calificaciones
- Mitologia JaponesaDocumento18 páginasMitologia JaponesaCristian Naula FaúndezAún no hay calificaciones
- Cementerio de San JavierDocumento2 páginasCementerio de San JavierEsteban García López100% (1)
- Comentarios PoemasDocumento10 páginasComentarios PoemassanchopanzaAún no hay calificaciones
- Actividad de MatemáticaDocumento8 páginasActividad de MatemáticaEsmeralda Muñoz Gómez100% (1)
- A ContracorrienteDocumento3 páginasA ContracorrienteSandra Elena Dermark BufiAún no hay calificaciones