También podría gustarte
- FORO 2 La Teoría EndosimbióticaDocumento2 páginasFORO 2 La Teoría Endosimbióticaeburbano12Aún no hay calificaciones
- Infografía LibroDocumento1 páginaInfografía LibroYAK BMAún no hay calificaciones
- Lynn MargulisDocumento1 páginaLynn MargulisLucía Álvarez EstebanAún no hay calificaciones
- Origen de Las Celulas EucariotasDocumento11 páginasOrigen de Las Celulas EucariotasJorge Oswaldo Toro JaimesAún no hay calificaciones
- 20170908210902Documento28 páginas20170908210902Nathaly iparraguirre rojas100% (2)
- Manual Ejercitacion XXVIOAB2017Documento156 páginasManual Ejercitacion XXVIOAB2017ceceort0% (1)
- El Origen de La Vida para Segundo Grado de SecundariaDocumento6 páginasEl Origen de La Vida para Segundo Grado de SecundariaEduardo Hurtado Perez100% (1)
- Hacia Un Sistema Natural de OrganismosDocumento1 páginaHacia Un Sistema Natural de OrganismosBriian Piipe Suarez CarvajalAún no hay calificaciones
- Biologia 3Documento152 páginasBiologia 3MARIANOAún no hay calificaciones
- Flujo de Materia en El EcosistemaDocumento5 páginasFlujo de Materia en El EcosistemaAlisson MariellAún no hay calificaciones
- Actividad Teórica 2 2014-00 MEHUDocumento3 páginasActividad Teórica 2 2014-00 MEHUOber SánchezAún no hay calificaciones
- AlgasDocumento3 páginasAlgaslissalfonsiaAún no hay calificaciones
- Teoria de La EvoluciónDocumento3 páginasTeoria de La EvoluciónBreiner Guancha100% (2)
- VertebradosDocumento14 páginasVertebradosDaryne De Gaytan Esquivel100% (1)
- Biologia ResumenDocumento15 páginasBiologia ResumenCarlaMolinaJerezAún no hay calificaciones
- El Origen de La Vida TeoriasDocumento4 páginasEl Origen de La Vida TeoriasmarcoAún no hay calificaciones
- Darwinismo y NeolamarckismoDocumento2 páginasDarwinismo y NeolamarckismoFacundo MotaAún no hay calificaciones
- Cuadernillo 1º Año Ciencias Naturales EES #13Documento45 páginasCuadernillo 1º Año Ciencias Naturales EES #13Leticia PerezAún no hay calificaciones
- Resumen para Faro QuímicaDocumento50 páginasResumen para Faro QuímicaGabriela JCAún no hay calificaciones
- Eugenio Tamayo - Pensamiento CríticoDocumento22 páginasEugenio Tamayo - Pensamiento CríticoAnonymous YFtULgD7Aún no hay calificaciones
- Trabajo Practico de Biología NM #8 - Glúcidos, Lípidos y ProteínasDocumento17 páginasTrabajo Practico de Biología NM #8 - Glúcidos, Lípidos y ProteínasJeremy Palma100% (2)
- Mod 5 - 4° Medio ElectivoDocumento19 páginasMod 5 - 4° Medio ElectivoSusan Sandoval Saavedra100% (1)
- Consumo de Drogas - PDF PDFDocumento6 páginasConsumo de Drogas - PDF PDFconstanzaAún no hay calificaciones
- CUADERNO DE EJERCICIOS N°5-Biología Mención 2016-Web PDFDocumento23 páginasCUADERNO DE EJERCICIOS N°5-Biología Mención 2016-Web PDFJose Manuel100% (1)
- TALLER DE BIOLOGIA - IcfesDocumento4 páginasTALLER DE BIOLOGIA - IcfesVICTOR PEÑAAún no hay calificaciones
- Respuesta Inmune en InvertebradosDocumento18 páginasRespuesta Inmune en InvertebradosWendyVanessaPulgarinOvalleAún no hay calificaciones
- CÉLULADocumento35 páginasCÉLULAMireya HerreraAún no hay calificaciones
- DemarDocumento1 páginaDemardatoval@gmail.comAún no hay calificaciones
- Biología SantyDocumento18 páginasBiología SantySantiago MattosAún no hay calificaciones
- Biología - (6 - GENÉTICA)Documento28 páginasBiología - (6 - GENÉTICA)CARLOS MARIO SARMIENTO MANRIQUEAún no hay calificaciones
- Guia de CnidariosDocumento3 páginasGuia de CnidariosandrewlAún no hay calificaciones
- Sistema EndocrinoDocumento7 páginasSistema EndocrinoNein OrtizAún no hay calificaciones
- Reinos de Los Seres Vivos para Segundo de SecundariaDocumento6 páginasReinos de Los Seres Vivos para Segundo de SecundariaJoel FuentesAún no hay calificaciones
- Guia 1 Teorias Origen de La VidaDocumento4 páginasGuia 1 Teorias Origen de La VidaMaria Jose Villarreal MenaAún no hay calificaciones
- Tema 7 GeneticaDocumento15 páginasTema 7 GeneticaWesley RobbinsAún no hay calificaciones
- Ser y PertenecerDocumento39 páginasSer y PertenecerBárbara Desántolo100% (1)
- Origen de La VidaDocumento15 páginasOrigen de La VidaAlex Rivera Ramos100% (1)
- Actividad 1 Origen de La VidaDocumento2 páginasActividad 1 Origen de La VidaRC100% (2)
- Pequeño Elogio Del Conocimiento - Federico Novelo UrdaniviaDocumento191 páginasPequeño Elogio Del Conocimiento - Federico Novelo UrdaniviaProfrFerAún no hay calificaciones
- 2 - Guia Pedagogica Biologia La Evolucion y Pruebas de La Evolucion Grado Noveno - 2021Documento3 páginas2 - Guia Pedagogica Biologia La Evolucion y Pruebas de La Evolucion Grado Noveno - 2021SaraAún no hay calificaciones
- Biología 1 Rosalino VázquezDocumento31 páginasBiología 1 Rosalino VázquezAna VázquezAún no hay calificaciones
- El Fenómeno Anómalo Del AguaDocumento3 páginasEl Fenómeno Anómalo Del AguaANA KAREN100% (1)
- CNIDARIOSDocumento9 páginasCNIDARIOSivanAún no hay calificaciones
- Guía Módulo BiologíaDocumento96 páginasGuía Módulo BiologíaAna Paula FariniAún no hay calificaciones
- Leyes de Mendel PDFDocumento13 páginasLeyes de Mendel PDFluz elena firigua pradaAún no hay calificaciones
- Teorias de EvolucionDocumento5 páginasTeorias de EvolucionJonasAún no hay calificaciones
- Teorías Del Origen de La VidaDocumento4 páginasTeorías Del Origen de La VidaII-A-LAURA ISIDORA AGUILAR VARGASAún no hay calificaciones
- Transporte en La MembranaDocumento10 páginasTransporte en La MembranaCRISTIAN MANUEL SAAVEDRA RODRIGUEZAún no hay calificaciones
- Historia de Los Reinos BiologicosDocumento18 páginasHistoria de Los Reinos BiologicosCompumuri MurilloAún no hay calificaciones
- Origen de La Vida BiologiaDocumento7 páginasOrigen de La Vida BiologiaJUAN RONDONAún no hay calificaciones
- Citologia Karp G Biologia Celular y Mole CAPITULO 1 A 5Documento213 páginasCitologia Karp G Biologia Celular y Mole CAPITULO 1 A 5Fatima AlvaradoAún no hay calificaciones
- Clase 3Documento34 páginasClase 3Carolina AravenaAún no hay calificaciones
- Proyecto de CatedraDocumento7 páginasProyecto de CatedraSamu JesúsAún no hay calificaciones
- Taller Origen de La Vida PDFDocumento5 páginasTaller Origen de La Vida PDFJULIETH VALENTINA GUERRERO MORENOAún no hay calificaciones
- Selectividad MembranasDocumento2 páginasSelectividad MembranasJesús Bustamante0% (1)
- Clasificcion de Los Seres Vivos - GrupoDocumento49 páginasClasificcion de Los Seres Vivos - GrupoSilver Tello FernándezAún no hay calificaciones
- Teoría de La EndosimbiosisDocumento3 páginasTeoría de La Endosimbiosis... marianaAún no hay calificaciones
- Teoría de La EndosimbiosisDocumento5 páginasTeoría de La EndosimbiosisCris HG100% (1)
- Teoría EndosimbiosisDocumento2 páginasTeoría EndosimbiosisLuisa FernandaAún no hay calificaciones
- Informe ObservaciónDocumento7 páginasInforme ObservaciónCarlos ChristianAún no hay calificaciones
- Guía de Diagnóstico, Tratamiento Nutricional y Rehabilitador de La Disfagia OrofaríngeaDocumento215 páginasGuía de Diagnóstico, Tratamiento Nutricional y Rehabilitador de La Disfagia OrofaríngeaFrancisco Watt50% (4)
- Ventajas y DesventajasDocumento2 páginasVentajas y DesventajasPerla Espino MarinAún no hay calificaciones
- Formato Guía para Elaborar Un PIAR (Tomado de Una I E)Documento6 páginasFormato Guía para Elaborar Un PIAR (Tomado de Una I E)Degnnys paola Monroy hernandezAún no hay calificaciones
- Switch Industrial Rapida SeleccionDocumento2 páginasSwitch Industrial Rapida SeleccionzerohuntercodeAún no hay calificaciones
- Analisis Protocolo SvaDocumento3 páginasAnalisis Protocolo SvaNeyda ChipanaAún no hay calificaciones
- Guia de Ejercicios 2023Documento27 páginasGuia de Ejercicios 2023Genaro Mendez CAún no hay calificaciones
- Didactik-Fichas DescriptivasDocumento7 páginasDidactik-Fichas DescriptivasIsaura MaldonadoAún no hay calificaciones
- VAG 1397 A GuionDocumento2 páginasVAG 1397 A GuionZelva ChileAún no hay calificaciones
- Formato para TR01Documento8 páginasFormato para TR01HENRY CACERESAún no hay calificaciones
- Desarmado de Baterías RecargablesDocumento3 páginasDesarmado de Baterías RecargablesRamiro SalazarAún no hay calificaciones
- Introduccion Del AceroDocumento56 páginasIntroduccion Del AceroFerni PelaezAún no hay calificaciones
- Guía EvaporadoresDocumento11 páginasGuía Evaporadoresmonica100% (1)
- Estados Financieros CESDocumento41 páginasEstados Financieros CESHarvi MartinezAún no hay calificaciones
- Actividad 18 Problemas de Tension Superficial - CompressDocumento8 páginasActividad 18 Problemas de Tension Superficial - CompresssenseytuproAún no hay calificaciones
- Ley SafcoDocumento3 páginasLey SafcoHijo De ConocimientoAún no hay calificaciones
- Lenguaje 1 Angela VelasquezDocumento11 páginasLenguaje 1 Angela VelasquezAngela VelasquezAún no hay calificaciones
- Powerflex RV KDocumento1 páginaPowerflex RV KPablo Hidalgo ValenzuelaAún no hay calificaciones
- ProcedimientoServicioApoyo-6278170 YAMILE LEALDocumento3 páginasProcedimientoServicioApoyo-6278170 YAMILE LEALMEGASERV POINT SASAún no hay calificaciones
- Informe Capilaridad, MECANICA DE FLUIDOSDocumento10 páginasInforme Capilaridad, MECANICA DE FLUIDOSAugusto Badel RubianoAún no hay calificaciones
- Fallas AceroDocumento33 páginasFallas AceroGustavo Adolfo100% (1)
- Control Quimico de La Placa BacterianaDocumento7 páginasControl Quimico de La Placa BacterianaSasha Nicole SantanderAún no hay calificaciones
- Planificación Digital DOCENTEDocumento41 páginasPlanificación Digital DOCENTEJeremy Brown100% (1)
- Anexo 1B. Argimon PJ. Estudios de CohortesDocumento10 páginasAnexo 1B. Argimon PJ. Estudios de CohortesJorge VargasAún no hay calificaciones
- CDU Abreviada 2016 PDFDocumento966 páginasCDU Abreviada 2016 PDFGabriela Salinas100% (1)
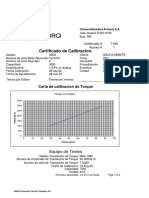
- ITEM 23 Cert - 1271872 - 30DX - 7189 Llave Torque RAD 30DXDocumento2 páginasITEM 23 Cert - 1271872 - 30DX - 7189 Llave Torque RAD 30DXJuanVasquezCarrascoAún no hay calificaciones
- Evaluacion Quimica 10 Periodo IIIDocumento3 páginasEvaluacion Quimica 10 Periodo IIIHeidy RölleAún no hay calificaciones
- MSDS 784-32 CatalizadorDocumento5 páginasMSDS 784-32 CatalizadorStrikererAún no hay calificaciones
- Actividad 2 Legislacion ComercialDocumento5 páginasActividad 2 Legislacion ComercialDiana LoaizaAún no hay calificaciones