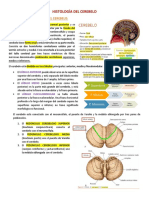
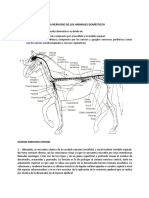
También podría gustarte
- Estructura Histológica de La Corteza CerebelosaDocumento19 páginasEstructura Histológica de La Corteza Cerebelosapatty41Aún no hay calificaciones
- Dialnet SubstanciaReticular 6293548Documento30 páginasDialnet SubstanciaReticular 6293548ALIDA MARINA AGUILAR GUTIERREZAún no hay calificaciones
- Barrera HematoencefalicaDocumento6 páginasBarrera HematoencefalicaVanessa Frisancho0% (1)
- NeurocienciasDocumento17 páginasNeurocienciasYoselin Valentina CastañoAún no hay calificaciones
- Bulbo RaquídeoDocumento7 páginasBulbo RaquídeoJoha VelasquezAún no hay calificaciones
- Ce RebeloDocumento14 páginasCe RebeloMaferAlonsoAún no hay calificaciones
- Anatomía e Histología de La RetinaDocumento10 páginasAnatomía e Histología de La RetinaReiniel Díaz Pérez100% (1)
- Tejido Nervioso y Sistema NerviosoDocumento16 páginasTejido Nervioso y Sistema Nerviosomsolorzano-1Aún no hay calificaciones
- Anatomía Del CerebeloDocumento3 páginasAnatomía Del CerebeloMila CcasaniAún no hay calificaciones
- Embiologia: Sistema NerviosoDocumento96 páginasEmbiologia: Sistema NerviosoBelen Muñoz RománAún no hay calificaciones
- Sistema nervioso: análisis histológico de la corteza cerebral, tálamo e hipocampo en caso clínicoDocumento7 páginasSistema nervioso: análisis histológico de la corteza cerebral, tálamo e hipocampo en caso clínicoJuan RodriguesAún no hay calificaciones
- Atlas de Histologà A Descriptiva 2012 PDFDocumento5 páginasAtlas de Histologà A Descriptiva 2012 PDFakuaAún no hay calificaciones
- Investigacion Del TimoDocumento4 páginasInvestigacion Del TimoYairely Perez GallegosAún no hay calificaciones
- Capítulo39 T2 2 ParteDocumento87 páginasCapítulo39 T2 2 ParteYasmira CerrónAún no hay calificaciones
- Cerebelo y Plexos CoroideosDocumento59 páginasCerebelo y Plexos CoroideosJackie RoblesAún no hay calificaciones
- MesencefaloDocumento17 páginasMesencefaloClinton anderson Luque mamaniAún no hay calificaciones
- Apunte Cerebelo 2010Documento14 páginasApunte Cerebelo 2010Marlyns Jovanna Valecillos CastilloAún no hay calificaciones
- Nervio FacialDocumento18 páginasNervio FacialAguus CabreraAún no hay calificaciones
- Sistema Nervioso ResumenDocumento6 páginasSistema Nervioso ResumenSofía ColzeraAún no hay calificaciones
- Anatomía Cerebro HistologíaDocumento6 páginasAnatomía Cerebro HistologíaMarlenMtz100% (1)
- Cerebelo. Neuro-2Documento15 páginasCerebelo. Neuro-2Gabriel DavalosAún no hay calificaciones
- Tejidos sanguíneo y nervioso: clasificación, estructura y funciónDocumento150 páginasTejidos sanguíneo y nervioso: clasificación, estructura y funciónLuis Enrique Vera CamposAún no hay calificaciones
- Función y anatomía del cerebeloDocumento15 páginasFunción y anatomía del cerebeloursula142100% (1)
- Disección cerebro resDocumento9 páginasDisección cerebro resLizette AngelAún no hay calificaciones
- Núcleo celular y cromatinaDocumento18 páginasNúcleo celular y cromatinaJesus Montemayor BeltranAún no hay calificaciones
- 1 MAT COMPL de SNC y CerebeloDocumento4 páginas1 MAT COMPL de SNC y CerebeloNelly DiazAún no hay calificaciones
- Histología Del CerebeloDocumento32 páginasHistología Del CerebeloanaAún no hay calificaciones
- Nucleo Celular - BiologiaDocumento17 páginasNucleo Celular - BiologiaDeisy Magdalena Morinigo UlderaAún no hay calificaciones
- Formación Reticular y Sistema LímbicoDocumento18 páginasFormación Reticular y Sistema LímbicoJorvelyn MartinezAún no hay calificaciones
- Características histológicas y función de los ganglios raquídeosDocumento2 páginasCaracterísticas histológicas y función de los ganglios raquídeosMarilsy Rondon de MejiasAún no hay calificaciones
- CEFALOCORDADOSDocumento20 páginasCEFALOCORDADOSJhon Contreras75% (4)
- Resumen de Cerebelo.Documento8 páginasResumen de Cerebelo.Daniel Velásquez CamposAún no hay calificaciones
- Nucleo CelularDocumento33 páginasNucleo Celularsirhecrulle100% (5)
- Generalidades Del CerebeloDocumento9 páginasGeneralidades Del CerebeloFernanda del Rocío Ramírez HernándezAún no hay calificaciones
- Nucleo CelularDocumento14 páginasNucleo Celularl1l2Aún no hay calificaciones
- Practica 5 - Anato 2Documento16 páginasPractica 5 - Anato 2Domenica Cuneo RomeroAún no hay calificaciones
- Cerebelo y DiencefaloDocumento9 páginasCerebelo y DiencefaloJose Achicahuala MamaniAún no hay calificaciones
- Diencéfalo y Telencéfalo 2013Documento16 páginasDiencéfalo y Telencéfalo 2013Felipe Andres MoyaAún no hay calificaciones
- Cerebelo Microanatomia IDocumento29 páginasCerebelo Microanatomia IAle MedranoAún no hay calificaciones
- Resumen de Histologia Del CerebeloDocumento15 páginasResumen de Histologia Del CerebeloLucia Saavedra LozadaAún no hay calificaciones
- Cerebelo: Estructura y Función del Órgano del Equilibrio y Coordinación MotoraDocumento8 páginasCerebelo: Estructura y Función del Órgano del Equilibrio y Coordinación MotoraAlfonsina AcostaAún no hay calificaciones
- Núcleo celular: estructura y funciónDocumento6 páginasNúcleo celular: estructura y funciónAnonymous VvMA4CTDAún no hay calificaciones
- Apuntes Tejido Nervioso PDFDocumento10 páginasApuntes Tejido Nervioso PDFFernandoAlioAún no hay calificaciones
- Tutoría El CerebeloDocumento81 páginasTutoría El CerebeloSebastián Alfredo Díaz RosalesAún no hay calificaciones
- Desarrollo Del Sistema NerviosoDocumento18 páginasDesarrollo Del Sistema NerviosoKaterine AvilaAún no hay calificaciones
- Neurofisiologia Del EncefaloDocumento31 páginasNeurofisiologia Del EncefaloJulian DavidAún no hay calificaciones
- Resumen Haines - CerebeloDocumento8 páginasResumen Haines - CerebeloMarisol Gómez ㋑Aún no hay calificaciones
- Capítulo de Atlas Citología e Histología 2005 Wolfgang PDFDocumento12 páginasCapítulo de Atlas Citología e Histología 2005 Wolfgang PDFHannie BertalotAún no hay calificaciones
- El Cerebelo (Modificado x1)Documento7 páginasEl Cerebelo (Modificado x1)albarranleivarossydarianaAún no hay calificaciones
- 23. Sistema Nervioso IIDocumento32 páginas23. Sistema Nervioso IImalubonfigliAún no hay calificaciones
- Sistema RenalDocumento37 páginasSistema RenalValentina Echeverry Nieto :3Aún no hay calificaciones
- Cápsula InternaDocumento5 páginasCápsula InternaAnthony Romero100% (2)
- Células del cerebroDocumento9 páginasCélulas del cerebroAngi Correa YanacAún no hay calificaciones
- Histología de la corteza cerebralDocumento8 páginasHistología de la corteza cerebralRaymundo PargaAún no hay calificaciones
- Núcleo Celular y MitosisDocumento8 páginasNúcleo Celular y MitosisNathalia GonzalezAún no hay calificaciones
- CerebroDocumento7 páginasCerebrobeautifulqueenAún no hay calificaciones
- Separata Sistema NerviosoDocumento17 páginasSeparata Sistema NerviosoVíctor Andres Mateo SanchezAún no hay calificaciones
- La unidad básica de la vida: la célulaDocumento6 páginasLa unidad básica de la vida: la célulaDL M LissetAún no hay calificaciones
- CAP 13 - Aprendizaje y MemoriaDocumento33 páginasCAP 13 - Aprendizaje y MemoriaSergio Miguel Polanco Santos0% (1)
- La plasticidad cerebral, neuronas espejo y poda neuronal en relación con el aprendizajeDocumento3 páginasLa plasticidad cerebral, neuronas espejo y poda neuronal en relación con el aprendizajeJoan Alexander Loli JaénAún no hay calificaciones
- Sistema NerviosoDocumento20 páginasSistema NerviosoJUANA KARLA RODRIGUEZ NAVAAún no hay calificaciones
- NeuropsicologiaDocumento36 páginasNeuropsicologiaJemi Vandevelde100% (3)
- Aparatos y sistemas del cuerpo humanoDocumento23 páginasAparatos y sistemas del cuerpo humanoRocio Galindo GarciaAún no hay calificaciones
- MetabolismoDocumento44 páginasMetabolismoSthephany LopezAún no hay calificaciones
- CoherenciaDocumento12 páginasCoherenciasilvinaAún no hay calificaciones
- Receptores sensitivos y transmisión de señalesDocumento9 páginasReceptores sensitivos y transmisión de señalesJORGE RODRIGO ALVA GALICIAAún no hay calificaciones
- Neuron CommunicationDocumento14 páginasNeuron CommunicationDan LópezAún no hay calificaciones
- Respuestas Guía N1 PDFDocumento7 páginasRespuestas Guía N1 PDFDamaris ApablazaAún no hay calificaciones
- Texto Divulgativo Bases Biológicas Del ComportamientoDocumento9 páginasTexto Divulgativo Bases Biológicas Del ComportamientoAlejandro ChisvertAún no hay calificaciones
- Neurofisiologia (Clase)Documento61 páginasNeurofisiologia (Clase)NaylaAún no hay calificaciones
- CyD - Receptores SensorialesDocumento7 páginasCyD - Receptores SensorialesFlorencia BallardiniAún no hay calificaciones
- Algologia TodoDocumento272 páginasAlgologia TodoZel Tzin Mkt100% (1)
- Neurotransmisores RYHHDocumento9 páginasNeurotransmisores RYHHDashly ReyesAún no hay calificaciones
- Libro de Psicologia BasicaDocumento12 páginasLibro de Psicologia BasicaHenry Abraham Ramos Gutierrez50% (2)
- Conceptos de NeuroplasticidadDocumento4 páginasConceptos de NeuroplasticidadDaniela Bermudez CalderonAún no hay calificaciones
- La Electroacupuntura Una Alternativa Contra Afecciones Médicas PDFDocumento4 páginasLa Electroacupuntura Una Alternativa Contra Afecciones Médicas PDFGustavo ParlantiAún no hay calificaciones
- Emoción: Relación Entre Emoción, Atención Y MemoriaDocumento7 páginasEmoción: Relación Entre Emoción, Atención Y MemoriaCentro de Enlace EducativoAún no hay calificaciones
- Organizacion de Nuestro CuerpoDocumento3 páginasOrganizacion de Nuestro CuerpoColegio Monte VerdeAún no hay calificaciones
- NEUROPLASTICIDADDocumento15 páginasNEUROPLASTICIDADJean Claude C. Hernandez100% (1)
- Linea Del Tiempo Psicobiologia y Teorias de La Evolucion y Preguntas Video Una Mente MaravillosaDocumento7 páginasLinea Del Tiempo Psicobiologia y Teorias de La Evolucion y Preguntas Video Una Mente MaravillosaJovana garcíaAún no hay calificaciones
- Trabajo de Investigacion EjemplosDocumento2 páginasTrabajo de Investigacion EjemplosKrisgaviAún no hay calificaciones
- Enfoque de Los Sistemas, El Ser Humano Como SistemaDocumento10 páginasEnfoque de Los Sistemas, El Ser Humano Como SistemaRhene MhendezzAún no hay calificaciones
- Sistema NerviosoDocumento5 páginasSistema NerviosodarlenisAún no hay calificaciones
- Guía Didáctica de Videos Sobre El Sistema Nervioso y Sus PatologíasDocumento7 páginasGuía Didáctica de Videos Sobre El Sistema Nervioso y Sus PatologíasGe MaAún no hay calificaciones
- Resumen FINAL-Parciales Neurofisiología. (1) (Autoguardado)Documento40 páginasResumen FINAL-Parciales Neurofisiología. (1) (Autoguardado)AlejoreAún no hay calificaciones
- Modelo Examen Escrito MFH IIDocumento6 páginasModelo Examen Escrito MFH IIJose Ibarra PerezAún no hay calificaciones
- Lo Publico y Lo Privado en El Ecuador: La Junta de Beneficencia de Guayaquil. 1880.1980Documento374 páginasLo Publico y Lo Privado en El Ecuador: La Junta de Beneficencia de Guayaquil. 1880.1980Patricia de la TorreAún no hay calificaciones