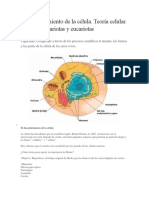
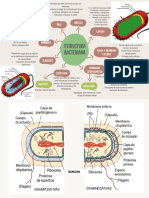
También podría gustarte
- Núcleo celular: estructura y funciónDocumento6 páginasNúcleo celular: estructura y funciónAnonymous VvMA4CTDAún no hay calificaciones
- Nucleo en BiologiaDocumento15 páginasNucleo en BiologiaAnnie BalderasAún no hay calificaciones
- Núcleo Celular - Wikipedia, La Enciclopedia LibreDocumento134 páginasNúcleo Celular - Wikipedia, La Enciclopedia LibreruizyannarithAún no hay calificaciones
- Nucleo Celular - BiologiaDocumento17 páginasNucleo Celular - BiologiaDeisy Magdalena Morinigo UlderaAún no hay calificaciones
- Nucleo CelularDocumento33 páginasNucleo Celularsirhecrulle100% (5)
- Núcleo Celular y MitosisDocumento8 páginasNúcleo Celular y MitosisNathalia GonzalezAún no hay calificaciones
- HISTORIADocumento2 páginasHISTORIAMileydi Centeno zubietaAún no hay calificaciones
- HistoriaTeoríaCelularDocumento29 páginasHistoriaTeoríaCelularRoberto EnriqueAún no hay calificaciones
- Reporte 1 Nucleo CelularDocumento5 páginasReporte 1 Nucleo CelularLuis Rey Cortes orijelAún no hay calificaciones
- Triptico Celula BiologiaDocumento3 páginasTriptico Celula BiologiaMIRNA75% (4)
- Teoría celular y sus postuladosDocumento38 páginasTeoría celular y sus postuladosMynor LucAún no hay calificaciones
- La Biología de La CélulaDocumento8 páginasLa Biología de La CélulaRossemary RivasAún no hay calificaciones
- Tema 2, Biologia, Devora ParedesDocumento13 páginasTema 2, Biologia, Devora ParedesFredy de JesusAún no hay calificaciones
- La CélulaDocumento27 páginasLa Célulamusicart30008394100% (2)
- Bloque II Biología Selectividad AndalucíaDocumento24 páginasBloque II Biología Selectividad Andalucíajulietttta100% (1)
- Documento Teoria CelularDocumento20 páginasDocumento Teoria CelularcalicoAún no hay calificaciones
- Tema 06 - Ciencias Básicas Tomo IIDocumento25 páginasTema 06 - Ciencias Básicas Tomo IIJuan FloresAún no hay calificaciones
- El Descubrimiento de La CélulaDocumento3 páginasEl Descubrimiento de La CélulaAnonymous Gjk3pBURAún no hay calificaciones
- M3 La Celula Unidad FundamentalDocumento24 páginasM3 La Celula Unidad FundamentalYesenia Toro CardozaAún no hay calificaciones
- La CélulaDocumento5 páginasLa CélulaalbertotreblaAún no hay calificaciones
- Taller CelulaDocumento66 páginasTaller CelulaLina VelezAún no hay calificaciones
- Trabajo Biologia GeneralDocumento29 páginasTrabajo Biologia GeneralJuan Camilo Quintero YustresAún no hay calificaciones
- Genética SeminarioDocumento7 páginasGenética SeminarioMayerling V. RiosAún no hay calificaciones
- Informe de Exposición NúcleoDocumento14 páginasInforme de Exposición Núcleomafer salazarAún no hay calificaciones
- UNIDAD III - La Celula Como La Unidad de Los Seres VivosDocumento60 páginasUNIDAD III - La Celula Como La Unidad de Los Seres VivosEmmanuel PaulinoAún no hay calificaciones
- Unidad 7 Cã©lulaDocumento29 páginasUnidad 7 Cã©lulaaleAún no hay calificaciones
- Biologia La CelulaDocumento10 páginasBiologia La CelulaINVERSIONES LucenaAún no hay calificaciones
- 5to PrimariaDocumento28 páginas5to PrimariaAnderson Rainiro Gordillo100% (1)
- Final de BotánicaDocumento136 páginasFinal de BotánicaNisla Yarileth De GraciaAún no hay calificaciones
- Teoria CelularDocumento7 páginasTeoria CelularLaelAún no hay calificaciones
- 5to. Año - BIOLOGÍA - Guía 3 - La Célula IDocumento6 páginas5to. Año - BIOLOGÍA - Guía 3 - La Célula IEdgar Feernando Martinez SanchezAún no hay calificaciones
- Libro de AnatomíaDocumento147 páginasLibro de AnatomíaRichard VeraAún no hay calificaciones
- Guia Nº13Documento5 páginasGuia Nº13Escuela Bajo PocoyanAún no hay calificaciones
- Biologia CelularDocumento26 páginasBiologia CelularJonathan Jose BravoAún no hay calificaciones
- Cuestionario de La CelulaDocumento3 páginasCuestionario de La CelulaLucyAún no hay calificaciones
- 5a - Teoria Celular. CélulasDocumento67 páginas5a - Teoria Celular. CélulasMauricio LiraAún no hay calificaciones
- 1 Biologa CelularDocumento33 páginas1 Biologa CelularKai Zuno0% (1)
- Núcleo celular y cromatinaDocumento18 páginasNúcleo celular y cromatinaJesus Montemayor BeltranAún no hay calificaciones
- Estructura y Funcion Celular IVDocumento29 páginasEstructura y Funcion Celular IVapi-399932263Aún no hay calificaciones
- 2 La Organización Celular PPT 1 PDFDocumento23 páginas2 La Organización Celular PPT 1 PDFLuis Fermini CastilloAún no hay calificaciones
- Guia Ciencias Naturales - PRIMER PERIODO SEXTODocumento14 páginasGuia Ciencias Naturales - PRIMER PERIODO SEXTOCarlos andres OcoroAún no hay calificaciones
- DR - ALBARRACIN-CELULA - PPT Filename UTF-8''DR ALBARRACIN-CELULADocumento94 páginasDR - ALBARRACIN-CELULA - PPT Filename UTF-8''DR ALBARRACIN-CELULAAndrea CondoAún no hay calificaciones
- Ley de CoulombDocumento10 páginasLey de CoulombSimonxd1316 Simonxd13Aún no hay calificaciones
- Nucleo Interfasico 1Documento26 páginasNucleo Interfasico 1José Luis Ramírez0% (2)
- La célula como unidad estructural y funcional de los seres vivosDocumento30 páginasLa célula como unidad estructural y funcional de los seres vivosEvelyn BertelloAún no hay calificaciones
- La célula: unidad fundamental de los seres vivosDocumento6 páginasLa célula: unidad fundamental de los seres vivosMax ParedesAún no hay calificaciones
- Sesión 12Documento8 páginasSesión 12Silvia Monica VillanuevaAún no hay calificaciones
- Biologia Informe ExposicionDocumento16 páginasBiologia Informe ExposicionAndrea CarvajalAún no hay calificaciones
- Ensayo GeneralidadesDocumento21 páginasEnsayo GeneralidadesRaúl Nolasco VelazquezAún no hay calificaciones
- La estructura y función de las células procariotas y eucariotasDocumento14 páginasLa estructura y función de las células procariotas y eucariotasAraceli NavAún no hay calificaciones
- Estudio de La CélulaDocumento26 páginasEstudio de La Célulahumberman100% (2)
- Presentación 4 La CélulaDocumento15 páginasPresentación 4 La CélulaAlejandro DelgadoAún no hay calificaciones
- Hojas de Aplicación para La WebDocumento2 páginasHojas de Aplicación para La WebMARIA FERNANDA CARRILLOAún no hay calificaciones
- Universidad Nacional Pedro Ruiz GalloDocumento22 páginasUniversidad Nacional Pedro Ruiz GalloBrain Jonatan SullonAún no hay calificaciones
- La estructura de la célula y sus componentesDocumento12 páginasLa estructura de la célula y sus componentesElizabeth AquinoAún no hay calificaciones
- Diferencias Estructurales de La Celula Animal y VegetalDocumento12 páginasDiferencias Estructurales de La Celula Animal y VegetalCarolina MtzAún no hay calificaciones
- Citología - I: La célula procariotaDocumento52 páginasCitología - I: La célula procariotaMarco PaucarAún no hay calificaciones
- Resum. Celulas Mendez 503Documento17 páginasResum. Celulas Mendez 503Mendez Angeles YairAún no hay calificaciones
- El núcleo celular: estructura, funciones y cromosomasDocumento42 páginasEl núcleo celular: estructura, funciones y cromosomasYen PaucarAún no hay calificaciones
- Bases Concurso XIIJuegosNacionalesDocumento5 páginasBases Concurso XIIJuegosNacionalesl1l2Aún no hay calificaciones
- Taller LinguisticaDocumento8 páginasTaller Linguistical1l2Aún no hay calificaciones
- Fisica WikiDocumento2 páginasFisica WikiMichael CobacangoAún no hay calificaciones
- Factores de riesgo y calidad de vida laboralDocumento84 páginasFactores de riesgo y calidad de vida laborall1l2Aún no hay calificaciones
- 13 Relaciones GramaticalesDocumento8 páginas13 Relaciones Gramaticalesl1l2Aún no hay calificaciones
- NanoalimentosDocumento4 páginasNanoalimentosl1l2Aún no hay calificaciones
- NanotecnologiaDocumento15 páginasNanotecnologiaAlan David LarreguiAún no hay calificaciones
- Huesos HumanosDocumento9 páginasHuesos Humanosl1l2Aún no hay calificaciones
- Globulos BlancosDocumento7 páginasGlobulos Blancosl1l2Aún no hay calificaciones
- HistoriaDocumento25 páginasHistoriaJHONATAN SANCHEZ MEGOAún no hay calificaciones
- Aparato RespiratorioDocumento10 páginasAparato RespiratorioDolores Ruiz SanchezAún no hay calificaciones
- La BiologiaDocumento22 páginasLa BiologiaElder GuzmanAún no hay calificaciones
- Nucleo CelularDocumento14 páginasNucleo Celularl1l2Aún no hay calificaciones
- Sistema DigestivoDocumento15 páginasSistema DigestivoKarla Ramirez100% (1)
- Sistema DigestivoDocumento15 páginasSistema DigestivoKarla Ramirez100% (1)
- NanoalimentosDocumento4 páginasNanoalimentosl1l2Aún no hay calificaciones
- Huesos HumanosDocumento9 páginasHuesos Humanosl1l2Aún no hay calificaciones
- NeuronasDocumento10 páginasNeuronasl1l2Aún no hay calificaciones
- AnatomiaDocumento7 páginasAnatomial1l2Aún no hay calificaciones
- NanotecnologiaDocumento15 páginasNanotecnologiaAlan David LarreguiAún no hay calificaciones
- Aparato RespiratorioDocumento10 páginasAparato RespiratorioDolores Ruiz SanchezAún no hay calificaciones
- Ejercicios FinancierosDocumento2 páginasEjercicios Financierosl1l2Aún no hay calificaciones
- 0fundam Calculo Financiero PDFDocumento33 páginas0fundam Calculo Financiero PDFWilder Sulca HuayanayAún no hay calificaciones
- FactorizacionDocumento18 páginasFactorizacionIvan Lopez MartinezAún no hay calificaciones
- Matematicas FinancierasDocumento4 páginasMatematicas Financierasl1l2Aún no hay calificaciones
- Acidos NucleicosDocumento2 páginasAcidos NucleicosGarmadilloAún no hay calificaciones
- Anticongelantes BiologicosDocumento12 páginasAnticongelantes BiologicosFelipaotao0% (1)
- Hla 2017 InmunoDocumento36 páginasHla 2017 InmunoMonica OrdinolaAún no hay calificaciones
- Evaluación Membrana y Transporte de MembranaDocumento6 páginasEvaluación Membrana y Transporte de MembranaCésar Humberto Villa Doyharcabal100% (1)
- Aspectos Metodologicos de La Expresion de Proteinas Recombinantes en E Coli PDFDocumento14 páginasAspectos Metodologicos de La Expresion de Proteinas Recombinantes en E Coli PDFJuan Jose OrtizAún no hay calificaciones
- Articulo - Usos de La ProteómicaDocumento4 páginasArticulo - Usos de La ProteómicaYazmin LopezAún no hay calificaciones
- Avances en biología molecular en los siglos XX y XXIDocumento9 páginasAvances en biología molecular en los siglos XX y XXINathanael De La Cruz AlmonteAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento13 páginasCadena Transportadora de Electronesagustin zarate cruz100% (1)
- Espectroscopía de absorción UV-Vis para la caracterización de biopolímerosDocumento25 páginasEspectroscopía de absorción UV-Vis para la caracterización de biopolímerosValentinaAlarcónAún no hay calificaciones
- La Célula Sus ComponentesDocumento8 páginasLa Célula Sus ComponentesBrayan CaoAún no hay calificaciones
- Respuestas Consolida 4 Semana 4Documento3 páginasRespuestas Consolida 4 Semana 4Ignacio González100% (1)
- Que Es El Adn en La EpigeneticaDocumento11 páginasQue Es El Adn en La EpigeneticaMaria Jose Perez100% (1)
- TRIPSINADocumento1 páginaTRIPSINAJorge HueichaAún no hay calificaciones
- Respiración CelularDocumento10 páginasRespiración Celularrodrigo paiva canoAún no hay calificaciones
- Ejercicio 2 - Unidad 2 - Ingrid SarmientoDocumento4 páginasEjercicio 2 - Unidad 2 - Ingrid SarmientoNatalia GaleanoAún no hay calificaciones
- Análisis de las tablas brasileñas de requerimientos nutricionales y perspectivas futurasDocumento5 páginasAnálisis de las tablas brasileñas de requerimientos nutricionales y perspectivas futurasluis carlosAún no hay calificaciones
- EjerciciosDocumento4 páginasEjerciciosAngelSaulSerranoAnguloAún no hay calificaciones
- Guía de Bioquimica (Aminoacidos, Proteinas y Enzimas)Documento8 páginasGuía de Bioquimica (Aminoacidos, Proteinas y Enzimas)Ines MuñozAún no hay calificaciones
- Metabolismo de Los Ácidos NucleicosDocumento16 páginasMetabolismo de Los Ácidos NucleicosGeo RosasAún no hay calificaciones
- Genoma Humano e Ingenieria GeneticaDocumento1 páginaGenoma Humano e Ingenieria GeneticaSharick GonzalezAún no hay calificaciones
- El Virus de La Gripe - FICHA TÉCNICADocumento5 páginasEl Virus de La Gripe - FICHA TÉCNICAVALERIA ALZATE MERIZALDEAún no hay calificaciones
- Examen de Biologia Andalucia 1Documento2 páginasExamen de Biologia Andalucia 1monografiasxAún no hay calificaciones
- Genética microbiana: genes, mutaciones y transferencia génicaDocumento66 páginasGenética microbiana: genes, mutaciones y transferencia génicaGloria Maza OsccoAún no hay calificaciones
- Proteinas FibrosasDocumento13 páginasProteinas FibrosasdianaAún no hay calificaciones
- Biocatalizadores y Referencias BibliograficasDocumento6 páginasBiocatalizadores y Referencias BibliograficasMaria Elizabeth M. Alberca0% (1)
- Clase 1b Ciclo Cero NobelDocumento80 páginasClase 1b Ciclo Cero NobelJorge Luis Plasencia CubaAún no hay calificaciones
- Estructura BacterianaDocumento2 páginasEstructura BacterianaFatima MartínezAún no hay calificaciones
- Sistematizacion y Transferencia 1.1Documento2 páginasSistematizacion y Transferencia 1.1Suarez MayAún no hay calificaciones
- ChoqueDocumento77 páginasChoqueMauricio Gomez100% (1)
- Ácidos NucléicosDocumento9 páginasÁcidos NucléicosCyber El CoyAún no hay calificaciones