También podría gustarte
- Libro Hematologia PregradoDocumento740 páginasLibro Hematologia PregradoRocio Sanchez Flores80% (5)
- LinfopoyesisDocumento12 páginasLinfopoyesisViianey Navii SpoockAún no hay calificaciones
- Sistema de Grupo Sanguíneo ABO 1 PDFDocumento20 páginasSistema de Grupo Sanguíneo ABO 1 PDFDavrob14Aún no hay calificaciones
- Biometria HemáticaDocumento5 páginasBiometria HemáticaBrenda ReyesAún no hay calificaciones
- Hepatitis CDocumento21 páginasHepatitis CJhon Jairo Medina DavisAún no hay calificaciones
- Capitulo 7 Robbins NeoplasiasDocumento45 páginasCapitulo 7 Robbins NeoplasiasMishelle Huamaní67% (3)
- Infeccion Por ChancroideDocumento24 páginasInfeccion Por ChancroideClau TomAún no hay calificaciones
- Hematología Básica - PinedaDocumento123 páginasHematología Básica - PinedaPaula LopezAún no hay calificaciones
- Valoración de Abdomen PDFDocumento21 páginasValoración de Abdomen PDFLaura OrosteguiAún no hay calificaciones
- Alteracion LeucocitariaDocumento17 páginasAlteracion Leucocitarialaura100% (1)
- Exploracion Fisica Del AbdomenDocumento78 páginasExploracion Fisica Del AbdomenValeria Moretto VegaAún no hay calificaciones
- HematopoyesisDocumento1 páginaHematopoyesisNayeli Fanola0% (1)
- Sistema de ComplementoDocumento1 páginaSistema de ComplementoRutis100% (3)
- Síndrome HemorragíparoDocumento2 páginasSíndrome HemorragíparoJasudeoAún no hay calificaciones
- Unidad 6. Sistema de Clasificacion de La Citologia CervicalDocumento26 páginasUnidad 6. Sistema de Clasificacion de La Citologia CervicalSantiago PérezAún no hay calificaciones
- Reparación de Tejidos - RobbinsDocumento14 páginasReparación de Tejidos - RobbinsGerson OrellanaAún no hay calificaciones
- Linfoma No HodgkinDocumento44 páginasLinfoma No HodgkinMENDOZA REYES LALY TATIANA100% (1)
- Cuestionario Examen Exploracion AbdominalDocumento5 páginasCuestionario Examen Exploracion AbdominalDereKunAún no hay calificaciones
- Escala de Bloom RichardsonDocumento3 páginasEscala de Bloom RichardsonEliannys LimongiAún no hay calificaciones
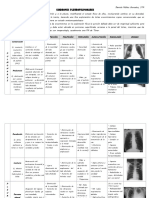
- SX PleuropulmonaresDocumento3 páginasSX PleuropulmonaresDanielaNuñezGonzalez100% (1)
- TrombofiliasDocumento27 páginasTrombofiliasAlejandro Jacob IturraAún no hay calificaciones
- Diagnostico Diferencial Dengue Chikungunya ZikaDocumento4 páginasDiagnostico Diferencial Dengue Chikungunya ZikaBere D LeonAún no hay calificaciones
- Reacción de ArthusDocumento6 páginasReacción de ArthusGloria María100% (2)
- Evaluación FonoaudiológicaDocumento2 páginasEvaluación FonoaudiológicaDaniela Díaz TorresAún no hay calificaciones
- Examen Anatomía y Fisiología AnimalDocumento3 páginasExamen Anatomía y Fisiología AnimalStefy MendezAún no hay calificaciones
- Drenaje Linfatico FinalDocumento26 páginasDrenaje Linfatico FinalValentina Alonso MeloAún no hay calificaciones
- Signo CervicotoracicoDocumento5 páginasSigno CervicotoracicokarenAún no hay calificaciones
- Artritis SépticaDocumento16 páginasArtritis SépticaChristian Rodriguez BermudezAún no hay calificaciones
- 4.2 ElectroAnalgesiaDocumento79 páginas4.2 ElectroAnalgesiaIrene JooAún no hay calificaciones
- Síndromes Deterioro Funciones Mentales Agudas y Crónicas 19-2Documento21 páginasSíndromes Deterioro Funciones Mentales Agudas y Crónicas 19-2Ruth Cobo Rosales100% (2)
- Fisiopatologia de Evc HemorragicoDocumento18 páginasFisiopatologia de Evc HemorragicovalfreAún no hay calificaciones
- Trombocitosis EsencialDocumento7 páginasTrombocitosis EsencialVania Rengifo100% (1)
- Anemias Hemolíticas Extracorpusculares o ExtrínsecasDocumento7 páginasAnemias Hemolíticas Extracorpusculares o ExtrínsecasAndreaRosales0% (1)
- MielopoyesisDocumento9 páginasMielopoyesisLeidy GonzabayAún no hay calificaciones
- Sindrome HemorragiparoDocumento50 páginasSindrome HemorragiparoMarco MoralesAún no hay calificaciones
- Historia Clinica 3Documento5 páginasHistoria Clinica 3Pedro Giovanni Alvarez GarciaAún no hay calificaciones
- Tipos de TóraxDocumento2 páginasTipos de TóraxGabriela Corrales CerdeiraAún no hay calificaciones
- Antigeno Prostatico EspecificoDocumento4 páginasAntigeno Prostatico EspecificoQUIMICO CLINICO WILLIANS SANCHEZ100% (2)
- Pelvis ÓseaDocumento6 páginasPelvis ÓseaAlex FSAún no hay calificaciones
- Enfermedad de Still Del Adulto (ESA)Documento14 páginasEnfermedad de Still Del Adulto (ESA)José Fernando Huanca CordovaAún no hay calificaciones
- Resumen de Hematolinfoide de RobbinsDocumento57 páginasResumen de Hematolinfoide de RobbinsMARIA JOSE TORRES ARTEAGAAún no hay calificaciones
- Historia Clínica UrológicaDocumento5 páginasHistoria Clínica UrológicaRaúl Valdez Vázquez67% (3)
- Resumen MurrayDocumento9 páginasResumen MurrayIchiNaru YhuSelAún no hay calificaciones
- Semiología Del Hemograma LabDocumento66 páginasSemiología Del Hemograma LabJose Garcia DumethAún no hay calificaciones
- BH, Quimica Sanguinea, EGODocumento11 páginasBH, Quimica Sanguinea, EGOivanvaloAún no hay calificaciones
- Sangre Valores Normales AdultosDocumento2 páginasSangre Valores Normales AdultosNora0% (1)
- Caso Clinico HemofiliaDocumento14 páginasCaso Clinico HemofiliaMaria TorresAún no hay calificaciones
- Histora Natural de Sindrome de Steve JohnsonDocumento3 páginasHistora Natural de Sindrome de Steve JohnsonDiany Sanchez67% (6)
- EstereotiposDocumento2 páginasEstereotiposMarthaMoralesAún no hay calificaciones
- Caso Clinico InmunologiaDocumento4 páginasCaso Clinico InmunologiaLauryMSierraAún no hay calificaciones
- 10 2020 - PATOLOGÍA DE GLÁNDULAS SALIVALES - Plantilla PPT UCSDocumento34 páginas10 2020 - PATOLOGÍA DE GLÁNDULAS SALIVALES - Plantilla PPT UCSGianmarco Rojas BellezaAún no hay calificaciones
- Endocarditis InfecciosaDocumento4 páginasEndocarditis InfecciosaJOSART ARATH CERNAS ORNELASAún no hay calificaciones
- HematopoyesisDocumento19 páginasHematopoyesisJocelyne Calderon PadillaAún no hay calificaciones
- Hematopoyesis MonseDocumento5 páginasHematopoyesis Monsejuan Cordova LaraAún no hay calificaciones
- Hematopoyesis ResumenDocumento4 páginasHematopoyesis ResumenErika Elizabeth Medina ToledoAún no hay calificaciones
- HEMATOPOYESISDocumento7 páginasHEMATOPOYESISAileen PresuelAún no hay calificaciones
- Articulo 2 HematologiaDocumento10 páginasArticulo 2 HematologiaRene Gtz SAún no hay calificaciones
- Celulas MadreDocumento9 páginasCelulas Madrearmanditocote94Aún no hay calificaciones
- Hematopoyesis AAyFADocumento15 páginasHematopoyesis AAyFAmarytayupegAún no hay calificaciones
- Aspectos Grales LmaDocumento11 páginasAspectos Grales LmaEduardo AnayaAún no hay calificaciones
- Actv 2 HematopoyesisDocumento7 páginasActv 2 HematopoyesisIngrid HernandezAún no hay calificaciones
- HematopoyesisDocumento12 páginasHematopoyesisKaren OlveraAún no hay calificaciones
- Hematopoyesis MonografiaDocumento9 páginasHematopoyesis MonografiaNicole100% (1)
- Taller AsincrónicoDocumento6 páginasTaller AsincrónicoJavier Buste AlvarezAún no hay calificaciones
- HematopoyesisDocumento7 páginasHematopoyesisHimitsu009Aún no hay calificaciones
- Células y Órganos Del Sistema InmuneDocumento17 páginasCélulas y Órganos Del Sistema InmuneNiponis ChunchunAún no hay calificaciones
- Hipertiroidismo y FAL ElevadaDocumento6 páginasHipertiroidismo y FAL ElevadasergioAún no hay calificaciones
- Unidad I Cuidados de Enfermería A La Mujer GestanteDocumento7 páginasUnidad I Cuidados de Enfermería A La Mujer GestanteDania HernándezAún no hay calificaciones
- Síndromes NeoplásicosDocumento33 páginasSíndromes NeoplásicosAlex FS100% (6)
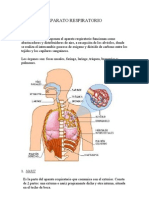
- Aparato RespiratorioDocumento9 páginasAparato RespiratoriobetenfAún no hay calificaciones
- Sondas VesicalesDocumento7 páginasSondas VesicalesYolanda Chazarra NavarroAún no hay calificaciones
- Resumo Urologia FinalDocumento97 páginasResumo Urologia FinalRaiane LimaAún no hay calificaciones
- Glomo Aórtico y CorazonDocumento7 páginasGlomo Aórtico y CorazonAnyle MendozaAún no hay calificaciones
- Fisiología RespiratoriaDocumento53 páginasFisiología RespiratoriaEnriqueCruz2Aún no hay calificaciones
- Fisiopatologia de La Falla Del Corazon Izquierdo (Documento14 páginasFisiopatologia de La Falla Del Corazon Izquierdo (Ruben SierraAún no hay calificaciones
- Laboratorio # 4 PDFDocumento6 páginasLaboratorio # 4 PDFJenny Potes MarbackAún no hay calificaciones
- TP Sistema Respiratorio y CirculatorioDocumento6 páginasTP Sistema Respiratorio y CirculatorioCamilooAún no hay calificaciones
- 10 Reproducción e Inseminación Artificial de Los Porcinos 31-Marzo-22Documento56 páginas10 Reproducción e Inseminación Artificial de Los Porcinos 31-Marzo-22moyra sanpiAún no hay calificaciones
- Aparato Urinario (Ok)Documento39 páginasAparato Urinario (Ok)Marco Luis Huaynalaya AlvaradoAún no hay calificaciones
- Inervacion Del Sistema Dentario y Las Estructuras PerimaxilaresDocumento2 páginasInervacion Del Sistema Dentario y Las Estructuras PerimaxilaresLuiggi Ruiz Sagal100% (2)
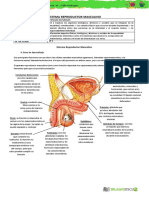
- Sistema Reproductor (Trab)Documento12 páginasSistema Reproductor (Trab)Vanessa Karina Gil RivasAún no hay calificaciones
- Linfocitos B. Patologias PDFDocumento6 páginasLinfocitos B. Patologias PDFcarl santiAún no hay calificaciones
- Anatomia Veterinaria - Sistema EndocrinoDocumento9 páginasAnatomia Veterinaria - Sistema EndocrinoDaniela Loranca ValdovinosAún no hay calificaciones
- Sistemas CibernéticosDocumento9 páginasSistemas CibernéticosCristina Fernández R100% (1)
- Embriolog A Del Sistema Genital FemeninoDocumento14 páginasEmbriolog A Del Sistema Genital Femeninoalemartin000Aún no hay calificaciones
- Trabajo Alimentacion 2Documento12 páginasTrabajo Alimentacion 2Esther Ballestero FernandezAún no hay calificaciones
- Cómo Funciona El OídoDocumento9 páginasCómo Funciona El OídoCele CelmanAún no hay calificaciones
- Sistema MasculinoDocumento4 páginasSistema MasculinoJuan Arnoldo Morales CarrascoAún no hay calificaciones
- Histología ApuntesDocumento37 páginasHistología ApuntesWinner Zapata NievesAún no hay calificaciones
- Términos Medicos Sistema GastrointestinalDocumento4 páginasTérminos Medicos Sistema GastrointestinalJohanna GómezAún no hay calificaciones